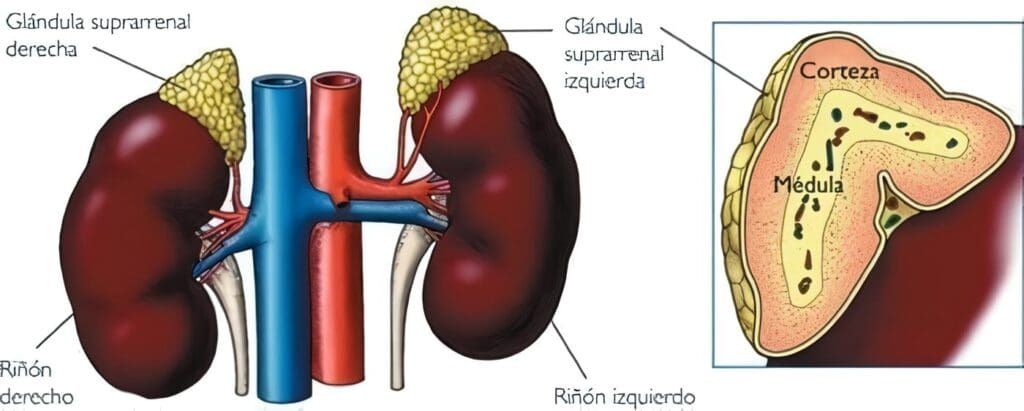
Fisiología de la Médula Suprarrenal
Autor: Alfredo F. Jácome-Roca, MD, FACP
Localizada a ambos lados al interior de las glándulas suprarrenales, la medula se encarga de sintetizar y segregar epinefrina (80%) y norepinefrina. La primera actúa básicamente como hormona, mientras que la segunda lo hace en el sistema nervioso como neurotransmisor, pero por tener niveles sanguíneos más altos que los de la epinefrina, también ejerce una función hormonal.
En las células blanco, estas catecolaminas se ligan a los receptores adrenérgicos donde inducen los mismos efectos que causa la estimulación de los nervios simpáticos. Las catecolaminas, compuestos fenólicos dihidroxilados, se sintetizan principalmente en las terminaciones de los nervios simpáticos, en la medula suprarrenal, el cerebro y en general en las células de las crestas neurales. Además de las dos mencionadas, está la dopamina.
FUNCIÓN ENDOCRINA DE LOS ADIPOCITOS
El tejido adiposo es uno de los tejidos más abundantes y representa alrededor del 15-20% del peso corporal del hombre y del 20-25% del peso corporal en mujeres. Debido a la baja densidad de los triglicéridos -moléculas grasas que los adipocitos guardan- y a su alto valor calórico, el tejido adiposo son muy eficientes en su principal función, almacenar energía para tiempos de ayuno, o de hibernación.
Cumple el tejido graso también una función aislante, que impide la pérdida del calor generado por las combustiones internas, protegiendo de la hipotermia. En los últimos años se ha destacado su función endocrina y metabólica, por la producción de una serie de hormonas -que actúan de manera endocrina, paracrina y autocrina- y que en conjunto se han llamado adipoquinas, que integran una red de señales que participan en la regulación de funciones en diversos tipos de células localizadas en órganos distantes, tales como hipotálamo, hígado, páncreas y músculo esquelético.
El tejido graso es además receptor de una serie de hormonas y proteínas que inducen cambios en él. El adipocito posee las enzimas que se requieren en la lipólisis y en la lipogénesis, procesos metabólicos finamente modulados por acción de hormonas, citocinas y otras moléculas implicadas en la regulación del metabolismo energético. Es capaz de modificar su tamaño hasta veinte veces su diámetro y varios cientos de veces su volumen.
Generalidades de la Fisiología de la Médula Suprarrenal
Localizada a ambos lados al interior de las glándulas suprarrenales, la medula se encarga de sintetizar y segregar epinefrina (80%) y norepinefrina. La primera actúa básicamente como hormona, mientras que la segunda lo hace en el sistema nervioso como neurotransmisor, pero por tener niveles sanguíneos más altos que los de la epinefrina, también ejerce una función hormonal.
En las células blanco, estas catecolaminas se ligan a los receptores adrenérgicos donde inducen los mismos efectos que causa la estimulación de los nervios simpáticos. Las catecolaminas, compuestos fenólicos dihidroxilados, se sintetizan principalmente en las terminaciones de los nervios simpáticos, en la medula suprarrenal, el cerebro y en general en las células de las crestas neurales. Además de las dos mencionadas, está la dopamina.
Anatomía e histología de la Médula Suprarrenal
La medula suprarrenal (que hace parte del sistema nervioso autónomo) tiene origen neuro-ectodérmico. Ocupa sólo el 10% del volumen de las glándulas suprarrenales y al corte presenta un color café oscuro. Las células poliédricas cromafinas de la medula se encuentran formando una red de cordones anastomóticos. A la microscopía electrónica se observan gránulos densos que contienen epinefrina, norepinefrina, ATP y varios neuropéptidos.
Sistema nervioso autónomo.
Este se divide en dos grandes grupos: la de la división tóraco-lumbar o “simpática” y la cráneo-sacra o “parasimpática”. La primera se origina de neuronas preganglionares situada en los segmentos medulares torácicos y lumbares superiores y la segunda, de neuronas preganglionares situadas en los núcleos de los pares craneanos motor ocular común (IV), facial (VII) y glosofaríngeo (IX), neumogástrico o vago (X) y en el 2º., 3º. Y 4º. segmentos sacros de la medula espinal.
Con la excepción de las vísceras inervadas por el vago, las demás reciben su inervación motora directamente de núcleos neuronales situados por fuera del sistema nervioso central. En el arco reflejo autónomo hay pues dos neuronas en relación con el ganglio mencionado, una preganglionar que comienza en el sistema nervioso central y otra postganglionar que comienza en el ganglio.
Así como hay reflejos somáticos en los cuales se mueve el músculo estriado (arco reflejo), también hay reflejos autónomos o vegetativos, ya que el músculo liso, el tejido glandular y el sistema de conducción cardiaca reciben innervación motora que al ser activa en forma refleja cambia el estado del órgano inervado. Las vísceras están pues bajo la influencia de esta subdivisión autónoma del sistema nervioso, a través de los neurotransmisores catecolamínicos o colinérgicos.
Hay dos tipos de neurotransmisores relacionados con el sistema nervioso autónomo: la acetilcolina y las catecolaminas epinefrina y norepinefrina. Tanto la norepinefrina como la acetilcolina se segregan y actúan en el sistema nervioso central. Las correspondientes al primero de estos neurotransmisores se llaman fibras adrenérgicas y las segundas, colinérgicas.
Entre las últimas están los nervios motores somáticos, todas las fibras autónomas preganglionares y todas las fibras postganglionares parasimpáticas. Entre las primeras están la mayoría de las fibras postganglionares simpáticas (con excepción de las que inervan las glándulas sudoríparas). Las fibras simpáticas que inervan estas glándulas liberan acetilcolina, al igual que las preganglionares simpáticas que inervan la medula suprarrenal.
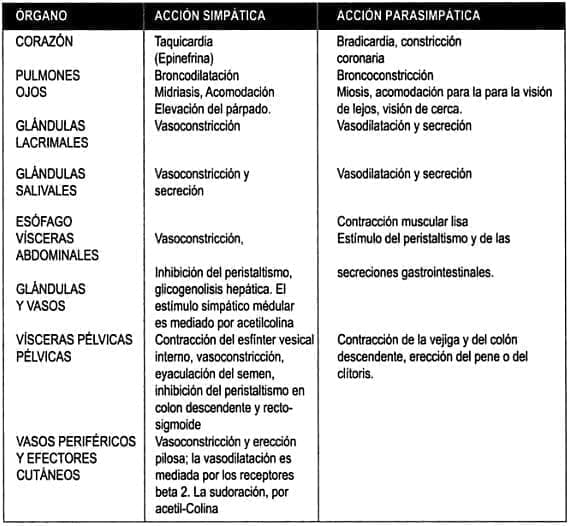
La mayoría de las vísceras bajo control autónomo (corazón, vasos coronarios, aparato digestivo, glándulas salivales, etc.) reciben doble innervación (simpática y parasimpàtica), mientras que otras (medula suprarrenal, glándulas sudoríparas, músculos pilomotores, muchos lechos vasculares que incluyen los de la piel y músculos) reciben sólo innervación simpática. En los sistemas inervados dualmente, los impulsos en las dos clases de nervios pueden actuar antagonísticamente. Las acciones de los nervios autónomos sobre las más importantes estructuras viscerales se resumen en la tabla 1.
Entre los ganglios paravertebrales y las vísceras pueden haber ganglios intermedios, con sinapsis preganglionares, como el ganglio celíaco, los mesentéricos superior e inferior, el ótico, el ciliar, el geniculado, etc.
Los núcleos hipotalámicos y el sistema límbico ejercen un influjo sobre el sistema nervioso autónomo. El estímulo de los centros superiores ante las situaciones de estrés produce reacciones de ataque o de huída. Dicho estímulo de los centros superiores produce cambios viscerales, vasculares y glandulares.
Tabla 1
Efectos neurovegetativos más conocidos
(Si los receptores beta causan relajación en un tejido, los colinérgicos producen contracción en el mismo tejido. Si los receptores alfa producen contracción en un tejido, los receptores colinérgicos regulan la relajación del mismo tejido).
Síntesis y metabolismo de las catecolaminas.- Provienen del aminoácido tirosina que es captado por las células cromafinas, transformándose en dihidroxifenilalanina (DOPA), por acción de una hidroxilasa, luego en Dopamina, por acción de una carboxilasa, enseguida en norepinefrina por otra hidroxilasa. Finalmente, esta a epinefrina, por acción de la feniletanolamina N-metil transferasa.
Esta última enzima sólo se expresa en la medula suprarrenal y en un ganglio simpático abdominal denominado órgano de Zuckerkland. La epinefrina y la norepinefrina se encuentran en una concentración de 2 a 4 mg por g de tejido medular. Una vez liberadas, ambas catecolaminas son rápidamente inactivadas en sangre (una infusión intravenosa sólo permite la recuperación del 4% en orina).
Las enzimas que intervienen en la deaminación y ortometilación son la catecol-O-metil-transferasa (COMT) y la monoaminooxidasa (MAO). Los metabolitos que se pueden medir en orina son la normetanefrina, la metafrina y el ácido vanililmandélico (VMA).
Secreción, liberación, transporte e interacción con los receptores
Las células medulares responden principalmente a la estimulación de los nervios esplácnicos (fibras simpáticas preganglionares) que liberan acetilcolina, la que genera la producción y liberación de las catecolaminas.
Si se seccionan dichos nervios, la actividad medular cesará. La hipoglicemia vacía los depósitos de epinefrina, mientras que la reserpina hace lo mismo, pero sólo con la norepinefrina. Además de la hipoglicemia, el ejercicio, una hemorragia y el estrés estimulan también la secreción de estos compuestos adrenérgicos. En la sangre se ligan muy laxamente con la albúmina, y tal vez con otras proteínas séricas.
Las respuestas adrenérgicas fueron clasificadas por Ahlquist en alfa y beta. Los alfa, median la contracción del músculo liso (excepto en el aparato digestivo) e inhiben secreciones celulares.
Los beta, median la relajación del músculo liso De esa época para acá ya se han identificado al menos nueve receptores, a saber: alfa-l (A, B y D). Alfa-2 (A, B y C). Beta-adrenérgicos (beta 1, 2 y 3) . Los receptores adrenérgicos son prototipos de las proteínas siete transmembrana, acoplados con las proteínas G que estimulan o inhiben señales intracelulares, que cuentan con cientos de miembros y que han sido descritos en la el capítulo sobre Generalidades.
Las varias clases de receptores adrenérgicos se acoplan con proteínas G cuyas sub-unidades alfa son también diferentes. Los beta-adrenérgicos 1 y 2 lo hacen con Gs para activar la adenil-ciclasa, aumentando el AMPc. En algunos tejidos, activan los canales de calcio. Los alfa 1- con Gq- para activar fosfolipasas como la C-beta, que producen IP3 y DAG, lo que aumenta la concentración de calcio en el citosol.
Como el aumento del calcio citosólico causa contracción del músculo liso, esta acción alfa-1 se traduciría en aumento de la tensión arterial por vasoconstricción y contracción del músculo liso intestinal, activando el peristaltismo.
Los alfa 2 se acoplan con Gi para inhibir adenilciclasa, disminuir la concentración de AMPc y en algunos tejidos regular los canales de potasio y calcio. Los receptores alfa 1, alfa 2 y beta 1 ligan efectivamente tanto epinefrina como norepinefrina, aumentando el calcio libre los primeros. Los receptores adrenérgicos beta dos sólo fijan epinefrina.
Hay alteraciones genéticas que pueden causar polimorfismos en ciertos locus de receptores beta adrenérgicos (un aminoácido es anormalmente reemplazado por otro) y así ocurre en casos de asma nocturna, mientras que un polimorfismo del receptor beta-3 puede llevar a los indios Pima norteamericanos a obesidad y aparición temprana de diabetes tipo 2, o en ciertos pacientes franceses, a obesidad mórbida.
Del conocimiento de estos sub-tipos se desprenden posibilidades terapéuticas. Por ejemplo, los receptores adrenérgicos alfa 1-A son los que en la próstata promueven la contracción del músculo liso, lo que haría que los actuales alfa-bloqueadores utilizados en esta patología, que pueden causar desagradables hipotensiones posturales, puedan ser reemplazados por nuevos agentes que no tengan este efecto colateral.
Como los receptores beta-3 promueven la lipólisis y la generación de calor en el tejido graso, nuevos agonistas de estos receptores resultarían útiles para el tratamiento de la obesidad. Los pacientes con falla cardiaca tienen una expresión aumentada de “receptores-proteínas G acopladas”, lo que aumenta la producción de cinasas y podría utilizarse en el desarrollo de drogas para tratar esta insuficiencia cardiaca.
Efectos fisiológicos de la Médula Suprarrenal
Como consecuencias funcionales de la liberación de catecolaminas, estas ayudan a lidiar el estrés. Entre los principales efectos podemos nombrar los siguientes:
1) Activación cardiovascular.
La frecuencia cardiaca y su fuerza de contracción aumentan, a través de su efecto sobre los receptores beta. A través de los alfa se produce una vasoconstricción generalizada que produce una elevación de la presión arterial por aumento de la resistencia periférica.
2) Suministro de energía.
La lipólisis suministra ácidos grasos para la producción de energía, conservando las reservas de glucosa. Pero también favorecen el desdoblamiento muscular de glicógeno, que suministra lactato para producción de glucosa por gluconeogénesis. El consumo de oxígeno y la producción de calor (metabolismo) se aumentan.
3) Mejora del estado de alerta.
Se estimula el sistema nervioso central, se dilatan los bronquíolos para respirar mejor y se dilatan las pupilas para ver mejor, particularmente cuando la luz ambiental es escasa.
4) Se llevan a cabo ciertos procesos no esenciales.
Como la inhibición de las secreciones y movimientos gastrointestinales.
Los efectos varían según la catecolamina comprometida. Por ejemplo la norepinefrina tiene mayor efecto vasopresor (por su efecto alfa adrenérgico), mientras que la epinefrina estimula el sistema nervioso, acelera el corazón, dilata los bronquios y activa la fosforilasa para producir glicógenolisis hepática y aumento de la glicemia de una manera más notoria que la norepinefrina (por su efecto betadrenérgico).
En cuanto a la lipólisis, constricción de vasos de la piel y estímulo respiratorio, ambas catecolaminas tienen igual efecto. Ciertas entidades clínicas alteran la sensibilidad a estas sustancias adrenérgicas, como el hipertiroidismo (que la aumenta) o el hipotiroidismo y la acidosis (que la disminuye).
Patología medular
La entidad clínica que se ve en la medula suprarrenal es una causa curable de hipertensión originada en un tumor frecuentemente benigno y a menudo unilateral, el Feocromocitoma. La sintomatología es paroxística y produce crisis adrenérgicas consistentes en hipertensión, taquicardia, empalidecimiento, sudoración y temblor entre otras manifestaciones.
A veces puede asociarse con otras entidades clínicas en la adenomatosis endocrina múltiple. El diagnóstico se hace por determinación urinaria de catecolaminas o de VMA, cuyas cifras se encuentran elevadas. Un TAC puede localizar el tumor. El tratamiento es quirúrgico, pero requiere cuidados especiales durante la extirpación.
Mecanismos que regulan la presión arterial.
Los centros vasomotores están en el bulbo raquídeo donde se encuentran dos discretas áreas, una caudal que media respuestas presoras y otra ventral, que regula respuestas depresoras. El aparato cardiovascular se encuentra bajo cierto nivel de estímulo de tipo simpático. Las vías descendentes (del tallo cerebral) median respuestas inhibitorias y excitatorias sobre el flujo simpático sobre corazón y vasos.
Por otro lado, el estímulo de los barorreceptores inhibe intensamente las neuronas de la medula rostral ventrolateral, mientras que el estímulo de los quimiorreceptores las excita. El núcleo del tracto solitario tiene una función integradora.
La distensión de la pared vascular causada por un aumento en la tensión arterial, produce un aumento de los impulsos aferentes a este último núcleo por parte de los barorreceptores, lo que disminuye la actividad simpática al tiempo que se aumenta la vagal, con vasodilatación arterial y venosa, disminución de la contractilidad miocárdica y bradicardia.
El tono vascular está controlado por una serie de factores, unos que producen vasoconstricción, otros vasodilatación, y otros alguno de los dos fenómenos.
Entre los primeros están el efecto alfa-adrenérgico de las catecolaminas, la angiotensina II y la aldosterona, la endotelina y la vasopresina. Entre los segundos, los péptidos natriuréticos, el óxido nítrico, el factor hiperpolarizante del endotelio, la adrenomedulina y la bradiquinina. Los eicosanoides como la prostaciclina son vasodilatadores mientras que el tromboxano es vasoconstrictor.
Sobre la volemia, el gasto cardiaco y la resistencia periférica inciden sustancias reguladoras del ión sodio como la aldosterona o el péptido auricular natriurético, vasoconstrictores periféricos como las catecolaminas y la angiotensina II, los barorreceptores y los centros vasomotores, que a su vez regulan la liberación simpática de catecolaminas o vagal de acetilcolina.
Nota histórica sobre la Fisiología de la Médula Suprarrenal
Bartholinus en 1651 describió la medula suprarrenal. El descubrimiento de la epinefrina se debe a Abel en1899, con estudios posteriores de Takamine y Abel en 1902. En los años 80, el británico James Black realizó una serie de trabajos sobre receptores de membrana, por lo que entre otras drogas descubrió el betabloqueador propanolol. Merced a sus investigaciones recibió posteriormente el Premio Nóbel.
Referencias seleccionadas
- Aristizábal D. Fisiopatología de la hipertensión arterial. En “Cardiología”, 1a. Ed. (R.Rozo, A. Merchán y cols. Eds.) Soc Col Cardiol Bogotá. 2000. Pp.365-373.
- Ahlquist RP. A study of the adrenotropic receptors. Am J Physiol 1948. 153: 586-600.
- Insel PA. Adrenergic receptors, evolving concepts and clinical implications. N Eng J Med 1996. 334: 580-585.
- Julius S. The evidence for a pathophysiologic significance of the sympathetic overactivity in hipertensión. Clin Exp Hypert 1996. 18:305-321.
- Melmon KL. The endocrinologic function of selected autacoids, catecholamines, acetylcholine, serotonin and histamine. In “Textbook of
- Endocrinology” (RH Williams, Editor). 6th Edition, WB Saunders, Philadelphia, 1981. Pp. 515-588.
- Varela E. Principios de biología molecular en el sistema cardiovascular. En “Cardiología”, 1ª.Ed. (R.Rozo, A. Merchàn y cols., Eds.) Soc Col Cardiol Bogotá. 2000. Pp.18-35.