Fisiología Endocrina
La fisiología endocrina estudia las funciones normales de las glándulas de secreción interna; al igual que la anatomía y otras disciplinas biológicas, puede ser comparada entre las diferentes especies de organismos multicelulares vivos y los seres humanos.
Las células endocrinas usualmente se acumulan en un órgano específico pero también se pueden encontrar diseminadas en otros aparatos, constituyendo el sistema endocrino difuso. Con el fin de coordinar la función celular en áreas como la reproducción, el crecimiento y desarrollo, la homeostasis o equilibro del medio interno, el ciclo sueño-estado de alerta y la regulación de la disponibilidad de energía, células especializadas liberan sustancias químicas a la sangre y a los líquidos tisulares, las que actúan como señales a otras células para controlar sus procesos.
Estos químicos se llaman “hormonas” (del griego “hormaos”, que significa excitar, estimular, revolver) y hacen parte de las moléculas que envían señales de una célula a otra
Historia.
En 2002 se cumplió el primer siglo del descubrimiento de la secretina, hecho por Bayliss y Starling en 1902. Estos fisiólogos, conocidos en investigación cardiovascular y digestiva, observaron –mientras estudiaban la digestión y absorción en un asa aislada de duodeno- que si el contenido duodenal era ácido, entonces el páncreas empezaba a segregar grandes volúmenes de jugo rico en bicarbonato.
Ya que dicha asa tenía irrigación más no inervación, ellos concluyeron que el estímulo se debía a una sustancia presente en la sangre, la que llamaron “secretina” y acuñaron el término “hormona”. Sorprende pensar que la secretina, de limitada utilidad en diagnóstico de la función pancreática, ahora esté de moda como una alternativa de tratamiento para ciertos casos de autismo, una enfermedad muy frecuente e incapacitante.
La teoría hormonal que siguió al descubrimiento de la secretina puso en jaque a la teoría neural, en boga en aquellos momentos. I.P. Pavlov –el de los reflejos condicionados, por lo que ganó el Nóbel de fisiología en 1904- postulaba que el funcionamiento de los órganos era debido al control nervioso. Pero ahora aparecía el concepto de hormona, el de los químicos que actuaban como reguladores biológicos a distancia.
Aunque por lo general las hormonas se refieren a la secreción de lo que los antiguos llamaban “órganos sin conducto (excretor)” –en contraposición a las glándulas exocrinas- curiosamente la secretina es producida por células que pertenecen al sistema neuroendocrino difuso.
Este concepto comenzó por la descripción en 1938 de unas células más bien pálidas, diseminadas en tejidos no endocrinos, hecha por Friedrich Feyrter. En 1969 A.G.Everson Pearse la histoquímica y ultra estructura de estas células productoras de pèptidos, las que denominó APUD.
Los primeros descubrimientos en la endocrinología fueron clínicos: la descripción por Addison de la Insuficiencia Suprarrenal, la diabetes –conocida desde la antigüedad- la Enfermedad de Basedow o el hipotiroidismo de Gull, el cretinismo, o incluso el “experimento” de Carlos Eduardo Brown-Sequard en 1879, quien se “rejuveneció” –al igual que su esposa- con la inyección de extracto testicular, generando gran interés en este campo. Sólo en el siglo XX se inició el estudio químico y fisiológico de las secreciones internas. La existencia del “medio interno” postulada en la centuria anterior por Claude Bernard obedeció sin embargo a la práctica de métodos experimentales.
En el campo de las hormonas, de los neurotransmisores y de los receptores se han premiado con el Nóbel a muchos investigadores, los que mencionaremos a lo largo de este libro. En este capítulo queremos especialmente mencionar a G.B. Elion, G.H. Hitchings y J. Black en cuanto a los receptores (1988); a R.A. Granit, H.K. Hartline, G. Wald (1969), D.H. Hubel, T.N. Wissel y R.W. Sperry en el campo de la fisiología y la bioquímica de la visión. A E.W. Sutherland (1971), quien descubrió el adenosìn-monofosfato cíclico (AMPc) como segundo mensajero en la transducción del mensaje hormonal, y a Martín Rodbell y Alfred G. Gilman (1994) que identificaron y purificaron la proteína G y que encontraron que era el transductor que ligaba el receptor hormonal con la amplificación de su respuesta.
Esta lista es bastante completa, pero no exhaustivas. Con frecuencia se descubren nuevos pèptidos y se conocen mejor sus genes de origen, por lo que se esperan muchas nuevas sustancias al dilucidarse por completo el genoma humano en el año 2003. La lista de citoquinas es también cada vez más larga.
Hay nuevos péptidos hormonales como el péptido liberador de prolactina, la adiponectina, otros factores de crecimiento como el IGF-2, etc. que se mencionarán más adelante.
¿Son hormonas todas las señales intercelulares?
Las primeras hormonas se descubrieron durante la investigación de enfermedades específicas; el déficit de insulina es causa de la diabetes, el de vasopresina, de la diabetes insípida, el de hormona tiroidea genera el cretinismo y el hipotiroidismo; su exceso, Enfermedad de Graves, el exceso o el déficit de hormona del crecimiento produce gigantismo o enanismo, etc. Ni el déficit de calcitonina ni el exceso de testosterona masculina o de progesterona femenina causan manifestaciones clínicas.
En algunas ocasiones, las enfermedades hormonales están causadas por fenómenos de resistencia en el receptor –caso de la diabetes tipo 2- a veces el exceso de secreciones internas proviene de formación ectòpica –como en los síndromes paraneoplàsicos- o hay situaciones en las que la hormona producida es estructuralmente anormal. Pero por regla general, tanto los excesos de producción hormonal (conocidos como enfermedades “hiper”) como las deficiencias (que resultan en patologías “hipo”), son causantes de manifestaciones clínicas.
En realidad el sistema nervioso y el hormonal actúan estrechamente, para constituir el sistema neuroendocrino. Las neuronas y sus axones envían estímulos eléctricos, que por despolarización de sus membranas mandan sus mensajes a gran velocidad, pero en la terminal nerviosa que hace parte de la sinapsis elabora químicos denominados neurotransmisores, que se fijarán a receptores localizados en la neurona post-sináptica, o en el órgano blanco directamente.
Esta integración neuroendocrina se observa no sólo en las células secretorias peptidèrgicas del hipotálamo, sino también en el sistema nervioso autónomo y en el control neural directo de algunas células endocrinas.
Los mensajes hormonales del sistema endocrino llegan a todas las células, pero sólo interactúan con aquellas que tienen receptores específicos que hacen que la célula responda a la señal dada por una hormona determinada. La endocrinología es entonces el estudio de las hormonas, de sus receptores y de las señales intracelulares producidas por la interacción de los dos primeros elementos.
El sistema inmune también está íntimamente relacionado con el neuroendocrino, ya que tiene una modulación neurohormonal para la producción linfocitaria de citoquinas, las que a su vez ejercen una influencia sobre las células endocrinas. Hay glosarios que definen a las interleucinas como hormonas segregadas por células inmunológicas (linfocitos, macrófagos y células dendríticas) que afectan otras células del sistema inmune al atraerlas, activarlas o inactivarlas.
El efecto de las citoquinas se ejerce por acción paracrina o autocrina. Sistemas similares existen, no sólo en vertebrados e invertebrados, sino incluso en organismos unicelulares; hay bacterias por ejemplo en las que se observan remedos más sencillos, que indican que los sistemas de comunicación se presentan temprano en la evolución.
Un complemento sobre esta sofisticada comunicación intercelular lo presentamos en el capítulo sobre el Sistema Neuro-inmuno-endocrino, que en el pasado se hubiera simplemente llamado sistema endocrino entèrico o en el mejor de los casos, sistema endocrino difuso. Tal vez la fisiología endocrina debiese llamarse de otra forma (algo así como “signalologìa intercelular”), a la manera como la radiología dio paso a la imaginología.
Las señales intercelulares son clásicamente endocrinas, pero también las hay autocrinas y paracrinas. Son autocrinas, cuando el producto celular sólo opera dentro del mismo citoplasma, o el químico segregado interactúa con el receptor de la membrana de esa misma célula; paracrinas, cuando entran a la circulación e interactúan a muy corta distancia, con receptores de células vecinas; se trataría por ejemplo del caso de las citoquinas que causan una respuesta inflamatoria local, o de la liberación de los neurotransmisores en los espacios sinápticos.
Se ha visto que hormonas que fundamentalmente actúan a distancia sobre otras células, pueden también servir de señales locales, tal como lo acabamos de mencionar. Pero además en otras ocasiones, la molécula química que da la señal se encuentra anclada a la membrana celular, y directamente –por medio de una conexión en muesca- interactúa con un receptor en otra célula inmediatamente vecina, dando una señal yuxtacrina.
La señal intercelular es intracrina cuando se produce la síntesis de esteroides localmente activos en tejidos de órganos blanco periféricos, como puede ser el caso de la producción prostática de dihidrotestosterona a partir de precursores suprarrenales inactivos allí como la DHEA y la androstenediona.
Estos términos –que parecen complicar el estudio de las señales intercelulares- han salido a flote gracias a la investigación en neuro-inmuno-endocrinología. En sentido estricto, no deberíamos llamar hormonas a las citoquinas, o a las moléculas ancladas a membranas, que se “cogen de la mano” a través de muescas, dando estímulos yuxtacrinos.
Tampoco a los neurotransmisores, a los medicamentos, ligandos que actúan por interacción con receptores o a los bloqueadores enzimáticos, a los agonistas y antagonistas de receptores, a muchos antiandrógenos y antiestrògenos, etc. Pero en sentido amplio, todas las células tienen actividad “endocrina”, por lo que con mayor frecuencia encontramos autores que llaman “hormonas” a estos químicos.
La serotonina liberada por la destrucción de plaquetas actúa localmente para producir vasoconstricción, mientras que la bradiquinina produce vasodilataciòn también local. Las hormonas neurovegetativas adrenales actúan sobre órganos a distancia cuando se liberan a la sangre, o localmente como neurotransmisores en las sinapsis; la dopamina es una hormona cuando actúa como PIH, pero es neurotransmisor en la generalidad de los casos, o precursor hormonal en el caso de la epinefrina.
Por extensión llamamos también hormonas a los análogos sintéticos de las hormonas naturales, sean pèptidos o esteroides. Pero básicamente aquí nos referiremos a las hormonas naturales, que son “agonistas”, es decir, a las moléculas que se ligan a un receptor e inducen todos los eventos post-receptor que generan un efecto biológico, con diferente potencia según el caso.
Por otro lado, los “antagonistas” son (usualmente medicamentos) que al ligarse al receptor, bloquean la fijación del agonista y así no generan la señal que produce los efectos intracelulares.
Hay quienes incluso consideran que las verdaderas hormonas son las hidrofìlicas, es decir los polipéptidos, mientras que las hidrofòbicas o liposolubles –como los esteroides y las hormonas tiroideas- se deberían llamar de manera diferente (pues no son mensajeros), por ejemplo, anacreones (un nombre futurible según Koeslag, mientras se aclaran bien ciertos aspectos de su fisiología).
También hay quienes hablan de parahormonas (como el gas carbónico y los hidrogeniones), productos finales del metabolismo celular que ejercen sus funciones en una amplia gama de blancos.
Estas comunicaciones intercelulares están afectadas por una serie de factores como la distancia (a medida que la sustancia – o la hormona- se difunde o es transportada desde el sitio de su producción, su concentración disminuye); la disponibilidad y densidad de los receptores celulares; el medio local (proteínas fijadoras, enzimas que degradan o alteran factores); la accesibilidad de la hormona (o de la droga que actúa como hormona) a la célula blanco potencial.
Dicho en otras palabras, la respuesta se aumenta o se disminuye cuando ocurre lo mismo con la fuerza de la señal, la densidad del receptor, o el proceso de inactivaciòn. Una hormona dada puede actuar con más fuerza en determinada célula blanco, porque el receptor tiene allí una gran afinidad por ella, pero actuar con poca fuerza en otra, pues el receptor de esta célula despliega escasa afinidad. Así como una hormona puede actuar en diferentes sitios, hay procesos fisiológicos que requieren la acción de varias hormonas, bien directamente o por un efecto permisivo que potencia la acción de la hormona primaria.
Clasificación de las hormonas.
Las hormonas pueden clasificarse en cuatro categorías según su estructura: 1) Pèptidos y proteínas, tales como las de la hipófisis o las pancreáticas .2) Esteroides, como las suprarrenales y gonadales. 3) Derivadas de aminoácidos, como las tiroideas o la epinefrina. 4) Derivados de ácidos grasos, los eicosanoides (prostaglandinas, prostaciclinas, leucotrienos y tromboxanos).
La tabla 1 nos muestra las diferentes hormonas, su estructura y su fuente principal de producción. Podemos anotar que no se han descubierto todas las hormonas que ocurren en la naturaleza, ni identificado todas sus funciones.
La terminología a veces confunde pues el nombre de la hormona se refiere a la primera acción que se le encontró (Vg. La hormona del crecimiento, nombre que tiene que ver con esa acción específica pero no con otras que posee, tales como la lipolìtica o la hiperglicemiante, o los estrógenos, que producen el estro, pero que también actúan sobre el hueso, la piel, los lípidos, etc.
Los factores de crecimiento, que no se mencionan en esa tabla, podrían considerarse hormonas naturales, y su importancia en el manejo de determinadas enfermedades no puede desestimarse. Muchos análogos de hormonas y varias antihormonas tienen un enorme valor terapéutico, particularmente en oncologìa.
Tabla 1 (De Kimball, modificada)
| TEJIDO GLANDULAR | HORMONA | SIGLA | ESTRUCTURA* | Segundo Mensajero |
| Adenohipófisis | Tirotrofina | TSH | Glicoproteína (201) | AMPc |
| H. Folículo estimulante | FSH | Glicoproteína (204) | AMPc | |
| Hormona Luteinizante | LH | Glicoproteína (204) | AMPc | |
| Prolactina | PRL | Proteína (198) | PK | |
| Hormona del crecimiento | HGH | Proteína (191) | PK | |
| Adreno-corticotrofina | ACTH | Proteína (39) | AMPc | |
| Neurohipófisis | Hormona-antidiurética | ADH | Polipéptido (9) | AMPc |
| Ocitocina | TRH | Polipéptido (9) | IP3 | |
| Tiro-relina | GnRH | Polipéptido (3) | ||
| Gonado-relina | GHRH | Polipéptido (10) | IP3 | |
| Somato-relina | CRH | Proteína (40) | AMPc | |
| Córtico-relina | GHR-IH | Proteína (41) | AMPc | |
| Somatostatina | Polipéptido (14) | |||
| Dopamina | DA | Derivado tirosínico | ||
| Pineal | Melatonina | Derivado del triptófano | ||
| Tiroides | Tiroxina | T4 | Derivado tirosínico | |
| Triyodo-tironina | T3 | Derivado tirosínico | ||
| Calcitonina | CT | Proteína (32) | AMPc | |
| Paratiroides | Hormona paratiroidea | PTH | Proteína (84) | AMPc |
| Corteza Suprarrenal | ||||
| Glucocorticoides | (Cortisol, cortisona, corticosterona) | Esteroides C 21 | ||
| Mineralocorticoides | (Aldosterona, DOCA) | Esteroides C 21 | ||
| Andrógenos Suprarrenales | (DHEA-S, androstenediona) | Esteroides C19 | ||
| Medula | Epinefrina | E | Derivado tirosí | IP3/AMPc |
| Suprarrenal | nico | |||
| Nor-epinefrina | NE | Derivado tirosí | IP3/AMPc | |
| Folículo Ovárico | Estradiol | E2 | Esteroides C18 | |
| Placenta | Estriol | E3 | Esteroides C18 | |
| Adiposito | Estrona | E1 | Esteroides C18 | |
| Cuerpo Lùteo, Placenta | Progesterona | P | Esteroide C 21 | |
| Trofoblasto, Placenta | Gonadotrofina coriónica | HCG | Glicoproteína (237) | |
| Testículos | Testosterona | T | Esteroide C 18 | |
| Islotes Langerhans | Insulina cèls.beta | Proteína (51) | PK | |
| Glucagòn cels.alfa | Proteína (29) | AMPc | ||
| Somatostatina cels.delta | Proteína (28) | |||
| Riñón | Eritropoyetina | EPO | Proteína (166) | PK |
| Calcitriol | Vit.D3 (1, alfa) | Derivado esteroide | ||
| Piel | Calciferol | Vit.D3 | Derivado esteroide | |
| Corazón | Péptido Auricular natriurètico | ANP | Proteìna (28,32) | GMPc |
| Estómago E Intestino | Gastrina | Polipéptido (14) | AMPc | |
| Secretina | Proteína (27) | |||
| Colecistoquinina | CCK | Polipéptido (8) | ||
| Somatostatina | Proteína (28) | |||
| Neuropèptido Y | Proteína (36) | NPY | ||
| Gonadorrelina | GnRH | Proteína(28) | IP3 | |
| PYY3-36 | Proteìna (34) | |||
| Factor insulinosímil de crecimiento-1 | IGF-1 | Proteína (70) | PK | |
| Hígado | Angiotensinògeno | Proteína | ||
| Trombopoyetina | Proteína (332) | |||
| Adipocito | Leptina | Proteína | ||
| · Los números entre paréntesis representan la cantidad de aminoácidos presentes en una proteína o en un polipéptido. · Los primeros trece órganos son los endocrinos clásicos y los últimos siete los endocrinos difusos. · Algunas hormonas producidas en el aparato digestivo se han llamado también cerebro-intestinales, pues se producen también en el sistema nervioso. · Varias de las hormonas aquí mencionadas se producen en células APUD. · PC= (Proteína cinasa, tirosina cinasa) · Los primeros 92 aminoácidos de los glicopèptidos FSH, LH, TSH y HCG son compartidos por ellos a través de la unidad alfa, que es idéntica en las cuatro hormonas. Los primeros 161 aminoácidos de la prolactina y de la hormona del crecimiento son idénticos, mientras que la GH y la somatomamotrofina coriònica contienen los mismos aminoácidos. La prohormona pro-opio-melanocortina contiene en su interior el ACTH y la beta-lipotropina, la cual a su vez es clivada a MSH y beta-endorfina. · Llamamos pèptidos a los compuestos con < 20 aminoàcidos y proteínas a los que contienen > 20. · Esta lista es bastante completa, pero no exhaustivas. Con frecuencia se descubren nuevos pèptidos y se conocen mejor sus genes de origen, por lo que se esperan muchas nuevas sustancias al dilucidarse por completo el genoma humano en el año 2003. La lista de citoquinas es también cada vez más larga. |
||||
Los polipéptidos y las proteínas son una forma química común de las hormonas. Ellas se sintetizan en el retículo endoplàsmico a partir de un precursor, se transfieren al aparato de Golgi y son empacadas en vesículas, antes de su liberación en forma de estallidos –la forma más común- que permite la rápida secreción de gran cantidad de hormona.
Las hormonas de naturaleza proteica se originan en grandes moléculas de “prohormona”, por ejemplo la Pro-opio-melanocortina, precursora del ACTH y de la MSH o también – encontrarse enclavadas en la secuencia de un precursor de mayor tamaño- son liberadas por clivajes proteolìticos. En la tabla 1 vemos que algunos son pèptidos con escasos tres aminoácidos mientras que otras son glicoproteìnas grandes, con múltiples subunidades, que son el producto de una traducción de un RNAm a un ribosoma.
Por regla general estas moléculas son hidrofìlicas –por lo que circulan sin necesidad de ligarse a proteínas circulantes-, tienen una vida media de pocos minutos e interactúan con receptores de membrana, originando la producción intracelular de un segundo mensajero.
Los péptidos hormonales recién sintetizados pueden liberarse de manera rápida – en respuesta a un estímulo secretorio corto- y, tanto los ya almacenados como los sintetizados de novo, se liberan lentamente –en respuesta a estímulos crónicos- produciendo niveles bajos estables de hormona circulante no ligada, cuyo tamaño relativamente grande previene la excreción renal rápida.
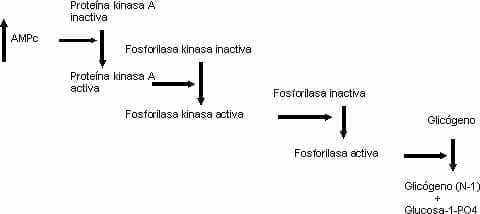
Cuadro 1. CASCADA DE EVENTOS INTRACELULARES DEL AMPc
Otra forma química común de las hormonas es la de los esteroides, lípidos que se derivan del colesterol y que tienen un núcleo de ciclo pentano-perhidrofenantreno. Son esteroides las hormonas gonadales y suprarrenales, así como también en cierta manera se consideran esteroides los calciferoles o vitaminas D.
En la parte interna de la membrana mitocondrial, el colesterol se transforma en pregnenolona, la que es sometida a múltiples transformaciones (esteroidogènesis) a ser enviada de ida y regreso al retículo endoplàsmico, a medida que nuevos precursores y hormonas se van formando.
La velocidad de la síntesis depende de la necesidad periférica del esteroide, ya que estos prácticamente no se guardan como depósito una vez sintetizadas. Al ser hidrofòbicas –pero liposolubles –se ligan de manera específica a ciertas proteínas (lo que determina su vida media y rapidez de eliminación) o inespecífica y con baja afinidad a proteínas como la albúmina.
Su liposolubilidad les permite atravesar la membrana de las células, e interactuar con receptores localizados en el núcleo, es decir, no necesitan de un segundo mensajero. En el hígado se inactivan, por ejemplo por glucuronizaciòn, y se eliminan por bilis y orina.
Las hormonas tiroideas y las catecolaminas se derivan del aminoácido tirosina, teniendo las primeras una vida media más o menos larga (de 3 a 10 días) mientras que las segundas circulan por unos pocos minutos. Otras hormonas-neurotransmisores que proceden de aminoácidos son la serotonina, que viene del triptòfano y que da lugar a la melatonina de la pineal, y la histamina, que se origina a partir del ácido glutámico.
Por otro lado los eicosanoides se derivan de ácidos grasos poliinsaturados, generalmente del ácido araquidònico presente en las membranas celulares, son transformadas y liberadas por la acción de lipasas permaneciendo activas por sólo unos segundos, y se clasifican en prostaglandinas, prostaciclinas, leucotrienos y tromboxanos.
Funciones hormonales.
Cada hormona puede tener funciones diferentes, aunque a menudo algunas son tal vez más importantes que otras. La acción puede ser directa –en los tejidos- o a través de la liberación de otras, como en el caso de las hormonas tróficas. En la tabla 2 podemos ver las más importantes.
Tabla 2 (De Raff, modificado)
Funciones generales que regulan las hormonas
| Reproducción | Ciclo menstrual, ovulación, espermatogènesis, embarazo y lactancia |
| Crecimiento y Desarrollo | Diferenciación sexual, características sexuales secundarias, aumento Masas ósea y muscular, velocidad del crecimiento linear |
| Homeostasis o Equilibrio del Medio Interno | Volumen extracelular, tensión arterial, equilibrio electrolítico y de Iones plasmáticos como el calcio, mantenimiento de las reservas Energéticas, como grasas, glicógeno, proteínas. |
| Suministro de Energía | Depósito, distribución y gasto de calorías, termogénesis |
| Conducta | Ingestión de agua y comida, libido y estado de ánimo. |
Control de la secreción hormonal.
Las hormonas hidrosolubles se pueden clasificar (según Koeslag) en tres diferentes categorías. A) Las hormonas que ejercen contrarregulaciòn de otras –control con riendas- o reguladoras del medio interno. B) Reguladoras balísticas, en las que los mensajes que envían no son luego confirmados (son de asa abierta) pues se generan en situaciones especiales o de emergencia. C) Mensajeros de relevo de hormonas, generalmente liposolubles, (que son de asa cerrada).
Entre las primeras, o que ejercen entre sí una contrarregulaciòn están la insulina y el glucagón (que regulan la glicemia), la PTH y la calcitonina (que regulan el calcio ionizado), la ADH y el ANP (que regulan osmolalidad, volumen sanguíneo, control del sodio), la EPO que controla la oxigenación de la hemoglobina, pero que no tiene una hormona contrarreguladora, y la gastrina y la secretina, que responden a la acidez gastroduodenal.
Reguladoras balísticas son la epinefrina, que de urgencia aumenta la glicemia y la tensión arterial, la ocitocina, que produce eyección làctea y contracción uterina, la prolactina, que regula producción de leche y la fertilidad, la CCK y las interleucinas. En cuanto a los mensajeros de relevo están las hormonas hipotalàmicas, las adenohipofisiarias u hormonas tròficas en general, la angiotensina II que regula la secreción de aldosterona y la PTH, que regula la l-alfa hidroxilaciòn renal de la vitamina D3, generando el calcitriol.
Las hormonas circulantes interactúan con las células blanco en mayor o menor grado de acuerdo a su concentración plasmática y a su afinidad por el receptor principalmente. La concentración depende de la velocidad de síntesis, liberación, degradación y eliminación.
Un mecanismo de control fisiológico común es la servo-regulación (que puede semejarse a la forma como funciona un calentador central, cuyo termostato actúa a una determinada temperatura ambiente, apagándolo o encendiéndolo. La retroalimentación (“feed-back”) más común en fisiología endocrina es la negativa en los diferentes ejes (hipotálamo-hipófisis con suprarrenales, gonadas o tiroides), que a veces puede ser positiva (estradiol y LH). Los circuitos pueden ser largos (hipófisis a glándula periférica), cortos (hipotálamo a hipófisis) o ultracortos.
La retroalimentación puede también operar por medio de la concentración plasmática de un sustrato; tal es el caso de la insulina y del glucagòn que se segregan en respuesta a un nivel dado de glicemia, o la parathormona y la calcitonina, que lo hacen de acuerdo a los niveles de calcio ionizado. Las hormonas se segregan de manera pulsátil, con pulsos cortos (de forma que logran evitar la regulación de receptores hacia abajo), o largos, de acuerdo a ritmos circadianos.
¿Cómo actúan las hormonas?
La forma como las hormonas actúan en las células blanco es por medio de activaciones enzimàticas o por modulación de la expresión genética, estimulando la trascripción de un grupo específico de genes. Esta actividad la logran por medio de la interacción, bien con receptores de membrana, o bien con receptores nucleares, según que la molécula sea hidrosoluble o liposoluble, lo que induce un cambio en la estructura del receptor.
La biología molecular y la ultraestructura nos muestran entonces que la morfología celular y la de sus componentes y organelas no permanecen estáticas, sino que cambia continuamente con los procesos fisiológicos. Aunque por razones didácticas se ha tratado de esquematizar la interacción de las moléculas señaladoras y sus receptores, estas acciones son complejas, tanto porque los receptores usualmente pertenecen a “superfamilias” con centenares de miembros, como porque los procesos comprometen innumerables proteínas, enzimas, iones y actividades genéticas.
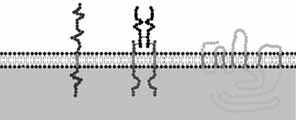
Como sabemos, las membranas celulares están compuestas de lípidos, proteínas y carbohidratos. La bicapa lipídica está integrada por fosfolìpidos con una cabeza polar hidrofìlica y una cola hidrofòbica ubicada en la parte central o media de la membrana; la composición varía en los aspectos internos y externos de la membrana celular lo que lleva a una asimetría: la parte externa contiene fosfatidil colina y esfingomielina (además de colesterol), mientras que la interna tiene fosfatidil serina, etanolamina e inositol.
Estos fosfolìpidos son móviles de manera lateral o transversa. Las proteínas de membrana son integrales (o sea que atraviesan la bicapa), o periféricas (bien en la parte externa o en la interna), y pueden ser móviles o estar fijas. Los carbohidratos se encuentran en la parte externa de la membrana plasmática, constituyendo glicoproteìnas y glicolìpidos, formando una capa conocida como glucocàliz, que sirve para fijar el calcio extracelular y de matriz de fijación para otras células.
El paso de sustancias a través de la membrana se hace por medio de mecanismos de difusión facilitada (simple o por medio de canales), o por transporte activo, cuando la difusión es mediada por moléculas portadoras. El oxígeno, el dióxido de carbono e incluso el agua, atraviesan la capa lipìdica con facilidad, pues son lipòfilas y sin carga. La glucosa, los aminoácidos y los iones, requieren de canales o de proteínas que ayudan al transporte hacia el interior de la célula.
Los transportes activos pueden ser primarios (como en el caso de la bomba de sodio/potasio) o secundarios; entre los últimos está el cotransporte, por una proteína energizada (Vg. Absorción de azúcares y aminoácidos en el aparato digestivo), o contra-transporte (intercambio de sodio por hidrógeno y por calcio).
Receptores de membrana y segundos mensajeros.
Los receptores en la membrana celular tienen regiones que contribuyen a tres dominios básicos, como son los extracelulares (o dominios que se fijan a los ligandos); los transmembrana (que incluyen aminoácidos hidrofòbicos que se insertan en la bicapa lipìdica y anclan el receptor en la membrana); muchos de ellos (pero no todos) atraviesan siete veces la membrana; y finalmente están los dominios intracelulares (colas o asas intracitoplàsmicas de los receptores), que son la región verdaderamente efectora hormonal, pues hay activación de proteínas cinasas, las que fosforilan proteínas, generalmente enzimas.
Estos receptores de membrana son de varias clases, siendo los más comunes los siete transmembrana acoplados a proteínas G (llamados 7TM o GPRC), proteínas que al ser activadas producen los segundos mensajeros, que actúan como señales intracelulares cuando la hormona no puede atravesar la membrana, como en el caso de las proteicas. Estos segundos mensajeros son: 1) el adenosìn monofosfato cíclico (AMPc). 2) El 1, 4,5-inositol trifosfato (IP3), el diacilglicerol (DAG)) y el calcio, que podríamos decir pertenecen al sistema fosfoinosìtidos-calcio. 3) El guanosìn monofosfato cíclico (GMPc). Una hormona puede utilizar diferentes segundos mensajeros y estos a su vez pueden ser producto de la interacción de los receptores con diferentes hormonas.
Como anteriormente decíamos, los receptores de membrana usualmente- pero no siempre- son los 7TM o GPCR, acoplados a proteínas G; pertenecen a una superfamilia que cuenta alrededor de mil miembros, entre los cuales están los adrenèrgicos, colinèrgicos, dopaminèrgicos, histaminèrgicos, receptores de prostaglandinas, y receptores de pèptidos del tipo vasopresina, ocitocina y angiotensina, de proteínas como glucagòn, FSH, LH y TSH, además de otros GPCRs en lengua, nariz y retina que pueden fijar moléculas con sabor, olor o luminosidad.
Este es el caso del receptor visual de luz – la rodopsina- cuya señal de transducción nos permite ver en la penumbra (visión de bastoncitos) o el color (visión de conos); diferentes rodopsinas absorben la luz de diversas longitudes de onda en forma máxima; la proteína G que allí se activa se llama transducina.
Estas proteínas G, fijadoras de guanina-nucleótido, son heterotrimèricas, y tienen tres sub-unidades: alfa (la que es activa, está ligada a guanosìn-difosfato o GDP y tiene actividad de GTPasa), beta (inactiva, de las que hay cinco conocidas) y gamma (diez conocidas); las sub-unidades a su vez tienen sub-familias.
Las sub-familias de la sub-unidades alfa son: la Gs (estimulante, precisamente la que es blanco de la toxina liberada por el Vibrio cholerae), que activa la adenil-ciclasa, aumenta el AMP cíclico que a su vez activa la proteína cinasa A; enseguida se produce la activación del factor CREB de trascripción (similar al HRE nuclear para los esteroides), que es la proteína fijadora del elemento de respuesta del cAMP; este CREB se liga a su elemento de respuesta en los promotores de los genes que responden a la hormona, lo que inicia la trascripción.
La secuencia de los eventos de la activación de las Gs comienza con la fijación del ligando al receptor, lo que al producir un cambio alostèrico o de conformación de la estructura terciaria de la proteína G, permite que la GDP ligada a la sub-unidad alfa sea desplazada por GTP; este, al ligarse, disocia la sub-unidad alfa, que se fija entonces a la adenil-ciclasa y la activa, lo que genera AMPc (Figura 1).
El GTP es hidrolizado por una GTPasa, lo que permite que la sub-unidad alfa deje la adenil-ciclasa y se reasocie con las beta y gama. Las Gs están asociadas con los receptores de hormonas tales como los beta-adrenèrgicos, ACTH, glucagòn, LH, FSH, TSH, PTH y vasopresina (acción renal). El caso clásico de la acción del segundo mensajero AMPc y la cascada subsiguiente es la activación de la fosforilasa hepática (por acción de la epinefrina y del glucagòn), que desdobla el glicógeno para producir glucosa 1 monofosfato, luego glucosa 6 fosfato que por acción de la glucosa 6 fosfatasa da lugar a glucosa, liberada a la sangre por el hígado. El AMPc se metaboliza por la acción de la fosfodiesterasa.
Otros receptores se acoplan con la proteína G-alfa-q (o Gq) que activa la fosfolipasa C beta, produciendo IP3 y DAG. El primero libera directamente calcio intracelular al citosol, cuando interactúa con su receptor, que es entonces fosforilado; el DAG, que activa la proteína cinasa C y puede actuar sinèrgicamente con el IP3, reduciendo la concentración de calcio necesaria para activar la cinasa.
El DAG y el calcio modulan la actividad de la proteína cinasa a través de la calmodulina; esta pertenece a una sub-clase de proteínas que fijan calcio, que activa una proteína fosfatasa, la calcineurina, enzima muy importante en la activaciòn de los linfocitos T, en la hipertrofia muscular y en la memoria retrógrada. Como el mantenimiento de niveles bajos de calcio citosòlicos es esencial para el buen funcionamiento celular, esto se logra entre otras cosas por el sistema calmodulina/ calcineurina que intervienen en actividades de fosforilación y defosforilaciòn de proteínas cinasas que son blanco del receptor de IP3.
La Gq se acopla a receptores como los de vasopresina (acción vascular), TRH, GnRH, y angiotensina II. Los canales de calcio (cuya compuerta en este caso sería abierta por ligandos intracelulares) se abren indirectamente por acción de los GCPR, a través de la formación de AMPc, GMPc, ATP o el mismo calcio.
Unos más (como el de la somatostatina, receptores alfa 2 adrenèrgicos y nor-adrenèrgicos) se acoplan con Gi para inhibir adenilciclasa y en algunos tejidos regular los canales de potasio y calcio. Las Go regulan los canales iónicos del sistema nervioso y las Gt (por transducina, que actúa sobre los bastoncitos de la retina), que hidrolizan el GMPc.
Otra forma importante de receptores es la de los tirosina cinasa, que tienen un dominio extracelular para unir los ligandos, un dominio transmembrana y otro citoplasmático, con actividad catalítica. Cuando se fijan hormonas la insulina, el IGF-1, y los factores de crecimiento epidérmico, el derivado de plaquetas y el fibroblàstico que utilizan este mecanismo, el receptor se autofosforila al producirse un cambio estructural. La cascada posterior produce la fosforilación de residuos tirosina en otras proteínas.
Como hemos mencionado anteriormente, hay hormonas que utilizan el ion calcio como segundo mensajero, mecanismo en el cual generalmente se encuentran involucrados otros segundos mensajeros como el DAG o el IP3 (fosfoinosìtidos), o bien en el aumento en el calcio citosòlico puede intervenir también la apertura de los canales de calcio en la membrana.