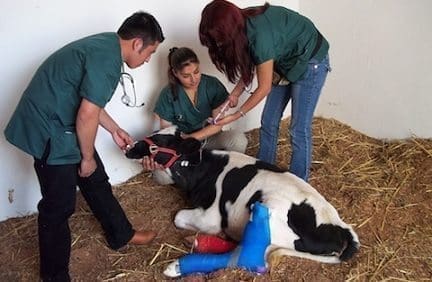
De las selenoproteínas las cuales han sido caracterizadas (Figura 1) cuatro son Glutarión peroxidasas. Estas enzirnas metabolizan peróxido de Hidrógeno y lipoperóxidos (Peróxidos de lípidos) en diferentes compartimentos de la célula. De esta manera el papel inicial del Se en los sistemas oxidativos puede ser explicado.
 Figura 1. Selenoproteínas en mamíferos
Figura 1. Selenoproteínas en mamíferos
Por ejemplo, la GSH-Px fosfolípido hidroxiperóxido, la cual protege la membrana celular de lipohidroperóxido, esta en probablemente la causa de las interacciones entre el Se y la deficiencia de vitamina E (Arthur y Beckett, 1994, citados por Arthur 1997).
Claves:
• (c GSH-Px) Glutation peroxidasa citosólica.
• (ph GSH-Px) Glutation peroxidasa fosfolípido hidroperóxido.
• (e GSH-Px) Glutation peroxidasa intracelular.
• (gi GSH-Px) Glutation peroxidasa gastrointestinal.
• (ID I, ID II, ID III) Tipos I, II y III de lodotironina deiodinasas. (Arthur, 1997).
La primera selenoproteína funcional en ser caracterizada en mamíferos fue la GSH -Px citosólica (c GSH -Px) (Rotruck et al., 1973, citaso pro Arthur, 1997), La pérdida de la actividad de la c GSH -Px permitirá la producción incrementada de especies de oxigeno reactivo, con el daño subsecuente a ácidos grasos insaturados en membranas y proteínas esenciales en toda la célula (Hoekstra, 1975, citado por Arthur, 1997). Junto con el descubrimeinto de la c GSH -Px del Se, aproximadamente el 30% del Se en la rata se halló en la proxidasa el resto es distribuido en otras proteínas (Belstein y Whanger, 1988, citado por Arthur, 1997). Las selenoproteínas lodotironina deiodinasas (ID I, II y III) regulan la conversasión de tiroxina (T4) a 3, 3, 5 – triyodotironina (T4) la hormona tiroidea activa o la hormona triyodotironina reversible (rT3) y la hormona tiroidea inactiva (Salvatore et al, 1995); 1996b; Larsen y Berry, 1995; Arthur et al., 1990, citados por Arthur, 1997). La ID tipo I (ID I) se encuentra en el hígado, riñón, cerebro, pituitaria y tejido adiposo café en rumiantes. La ID tipo II (ID II) se encuentra en cerebro, y pituitaria de las especies examinadas y tejido adiposo café en humanos.
La ID II catiliza la conversión de T4 a T3. La ID III convierte de T4 a T3 y T3 a diyodotironina, se encuentra en cerebro, piel y placenta y su función es desactivar las hormonas tiroideas (Larsen y Berry 1995, citados por Arthur, 1997). Un efecto característico de la deficiencia de Se en las hormonas tiroideas plasmáticas causado por los descensos en la actividad de ID I es un incremento marcado (superior al doble) en la concentración de T4 y un descenso más pequeño en la concentración de T3 (Beckett et al., 1987, 1989, citados por Arthur, 1997). Sin embargo, en deficiencias de Se prolongadas, las concentraciones de la hormona tiroidea plasmática pueden retornar a los niveles control a pesar del gran descenso en la actividad de la ID I en el hígado y riñón (Mitchell et all.), 1996, citados por Arthur, 1997).
| El esperarma de los animales con deficiencia de se tiene pobre motilidad de características del desarrollo anormal de la cola del esperma. La deficiencia de Se no solamente precipitó este problema en verracos, sino que el esperama deforme fue menos efectivo en la subsecuente fertilización del Oocito ovulado. |
| Los lechones son particularmente susceptibles a problemas asociados a una deficiencia de la vitamina E y Se. Al crecer el lechón de la E y el Selenio se acumulan en los tejidos esto es si la dieta tiene buenos niveles de estos dos elementos. al destete sin embargo, se presenta un decrecimiento del alfa-tocoferol y del Se sanguíneo, y puede ser relacionado con el síndrome de muerte súbita. |
Otras selenoproteínas que se han identificado son la selenoproteína P, selenoproteína W, tioredoxin reductasa, proteínas selenoligadas (58,, 56 y 14 kda), selenoproteína de la cápsula espermática y una selenoproteína en células epiteliales glandulares de la próstata y ratones. La selenoproteína y da razón de 60 – 80% del Se del plasma (Burk y Hill, 1994, citados por Arthur, 1997). La función de la selenoproteína P permanece incierta, aunque se pensó originalmente que tenía una función de transporte, distribuyendo el Se a los diferentes órganos por la circulación. La síntesis preferencial de selenoproteína P en ratas deficientes en Se, asociado con una protección incrementada contra el Dicuat, ha llevado a la propuesta de una actividad antioxidante o estabilizante para la proteína de la membrana celular (Burk et al., 1995 citados por Arthur, 1997).
La selenoproteína W se manifiesta en músculo esquelético con una subunidad molecular con peso de 9,5 Kda. La función de la selenoproteína W no se ha identificado, se postula que tiene propiedades antioxidantes. La pérdida de esta selenoproteína se asocia con la enfermedad del músculo blanco en ovejas, combinadas las deficiencias de Se y vitamina E (Levander y Whanger., 1996, citados por Arthur, 1997)-
La tioredoxin reductasa fue purificada de un grupo de células cancerígenas de pulmón humano, contiene Se como una selenocisteina en el penúltimo AA de terminación C de la proteína. (Nicol y Arthur, sin publicar, citados por Arthur, 1997.
Una selenoproteína en la cápsula espermática ha sido identificada, la cual es consistente con el papel del Se en el mantenimiento de la fertilidad normal (Cataldo et al, 1996, citados por Arthur, 1997). La selenoproteína es el mayor componente de la cápsula espermática y se ha postulado que tiene una función estructural (Kalcklosh et al., 1995, citados por Arthur., 1997).
Poco se conoce sobre las funciones del Se en las proteínas ligadas 14.56, 58 Kda. Para la instancia la proteína Kda está ligada a un ácido graso y es incierto si el Se ligado tiene una función fisiológica (Sinha et al., 1993, citados por Arthur. 1997).
Efectos de la vitamina E y Selenio sobre la respuesta inmune
Tanto la vitamina E como el Se son importantes en el mejoramiento de la integridad celular, e importantes medios para proporcional inmunidad innata al animal (Figura 2).
 Figura 2. Inmunidad innata
Figura 2. Inmunidad innata
La vitamina E es incorporada a la matriz lipoproteíca de la pared celular protegiendo así contra el daño oxidativo.
El Selenio en la forma de GSH-Px, es una de las varias enzimas que actúan desde el citoplasma, protegiendo el medio ambiente celular de daños oxidativos de procesos metabólicos.
Como la vitamina E y el Selenio proveen una respuesta inmune vía integridad celular y una estimación del sistema humoral y celular, la respuesta inmunogénica a la enfermedad es incrementada.
Tiege et al. (1982), citado por Combs y Combs (1986), evaluaron la respuesta de cerdos alimentados con Selenio 0,2 mg o 200 U.I. de vitamina E por día, y desafiando con Serpulina Hyodisenteria/(Disentería porcina). Tanto la vitamina E como el Selenio disminuyeron la mortalidad porcina y los niveles de S, Hyodisenteria/recuperados de las heces. Los signos clínicos de disentería fueron menos severos con suplementación de Selenio.
Los lechones son particularmente susceptibles a problemas asociados a una deficiencia de vitamina E y Selenio. Al crecer el lechón la vitamina E y el Selenio se acumulan en los tejidos esto es si la dieta tiene buenos niveles de estos dos elementos. Al destete, sin embargo, se presenta un decremento del alfa – tocoferol y del Se sanguíneo, y puede ser relacionado con el síndrome de muerte súbita; es por esto que la suplementación en esta época es bastante importante (Combs y Combs., 1S86).
Los efectos de la deficiencia de Se sobre el sistema inmune y sobre las propiedades tóxicas de los virus requiere de una posterior investigación para clasificar la investigación para clasificar la causa subyacente. La deficiencia de Se empeora varios aspectos del orden immunitario de la función del neutrófilo para la resistencia a la infección (Levander et al., 1995; Turner y Flinch., 1991, Boyne y Arthur, 1986, citados por Arthur, 1997). La actividad disminuida de la Cgsh-Px podría llevar a los peróxidos maten al neutrófilo tan pronto como la célula esté fagocitando (Boyne y Arthur., 1986; citados por Arthur., 1986; citados por Arthur, 1997). Sin embargo, es menos fácil relatar otros cambios inmunes en la deficiencia de Se, tales como la producción deteriorada de anticuerpos y la baja actividad de la c GSH-Px.
Del mismo modo como el deterioro del sistema inmune del hospedero, la deficiencia de Se puede efectuar cambios en los microorganismos infecciosos. Un Coxsakievirus B3 no cardiotóxico en ratones llega a ser cardiotóxico luego que pasan a través de una deficiencia de Selenio. Este cambio en el fenotipo se asocia con cambios específicos en las bases del genoma viral, aunque el mecanismo por el cual esto ocurre se desconoce (Berck., 1996; citado por Arthur., 1997).