Diana García de Olarte MD.1, Carlos Julio Montoya Guarín MD., MSc.2
1. Pediatra, DSc, Profesor Titular Universidad de Antioquia.
2. Profesor del Centro de Investigaciones Médicas.
Grupo Patogénesis de las Inmunodeficiencias Primarias, Facultad de Medicina, Universidad de Antioquia.
Desde hace 20 años existen en el comercio preparaciones seguras de gammaglobulina G intravenosa, lo cual implicó un gran avance en el tratamiento de las inmunodeficiencias primarias.
Adicionalmente, aparecen informes clínicos que evidencian como puede ser modulada la respuesta inmune, con la administración de grandes cantidades de gammaglobulina intravenosa, y los mecanismos responsables de esa regulación siguen su investigación.
Por estos hallazgos clínicos, cada día emergen nuevas indicaciones para la terapia con gammaglobulina intravenosa.
Palabras claves: gammaglobulina intravenosa, inmunomodulación.
Abstract
Since 20 years ago safe preparations of immune globulin suitable for intravenous use became available.
It was a great advance in the treatment of primary immunodeficiency diseases; furthermore, increasingly clinical evidences have been reported about that immunologic reactions can also be modified by the intravenous administration of large amounts of gammaglobulin, and the mechanisms that may be responsible for such immunoregulation are under investigation.
Several indications for intravenous gammaglobulin therapy are emerging due to those clinical findings.
Key words: intravenous gammaglobulin, immunomodulation.
Introducción
Las inmunoglobulinas (Igs) son glicoproteínas producidas por las células plasmáticas. Los cinco isotipos de inmunoglobulinas: M, D, G, A, E, son las principales moléculas efectoras del sistema inmune humoral.
El 75% de las Igs séricas está representado por la IgG y provee la mayor parte de la actividad de anticuerpos (acs) contra diferentes microorganismos infectantes de diseminación hematógena: bacterias, virus, parásitos y hongos. Fija complemento (C’) y facilita la mayoría de las reacciones antígeno-anticuerpo (ag-ac).
Las preparaciones de Igs a partir de sangre humana se han utilizado en medicina clínica desde 1952 para tratar enfermedades por deficiencia inmune.
En los últimos años se ha dispuesto de diversas preparaciones de IgG para aplicación intravenosa (IgGIV). Las que se obtienen en el mercado contienen 3, 5 ó 6% de proteínas; más del 90% de la proteína es IgG y sólo tienen trazas de IgA e IgM.
En la mayoría de estos productos, la composición de los isotipos de IgG es similar a la que se encuentra en el suero normal (1).
La IgGIV está compuesta en el 95% por IgG monomérica, con pequeñas cantidades de IgG dimérica o agregada, sin embargo, después de la infusión, algunos monómeros de IgG se convierten en dímeros o complejos con ags circulantes encontrados en el paciente.
Los monómeros se unen frecuentemente a los receptores FcgRI (receptor para la fracción Fc de las inmunoglobulinas), localizados en los monocitos, mientras que los receptores FcgRII y FcgRIII están en los monocitos, neutrófilos (PMN) y otras células e interactúan con la IgG polimérica o los complejos inmunes.
Si la molécula de ac está unida al ag, se generan cambios de configuración en la fracción Fc (Fracción constante), que facilitan la unión al FcgR.
La unión específica de un ag con una molécula individual de IgG:
Es conferida por el arreglo estérico tridimensional inherente a las secuencias de aminoácidos (aa) de las regiones variables (Fab: sitio de unión a los ags o paratope) de las cadenas tanto pesadas como livianas de las moléculas. Esta porción de la IgG se denomina el determinante idiotípico.
Solo un determinante antigénico se adapta bien con este arreglo. Aparte de la región del idiotipo, el resto de la molécula de IgG es relativamente constante y se designa Fc.
Al encuentro con un ag, las células T (CT) inductoras y luego las células B (CB), reaccionan a cada determinante antigénico sintetizando IgG; estos acs se unen al ag, pero una familia específica de CT y B inductoras están presentes en el repertorio de reconocimiento del ag, las cuales pueden reconocer el determinante idiotípico de la IgG, lo que lleva a la producción de acs-antiidiotípicos.
El antiidiotipo tiene determinantes antigénicos (idiotipos), que pueden ser idénticos al ag que inició la reacción (2) (figura 1).
Los acs antiidiotípicos provocan su propia respuesta antiidiotípica y por tanto, muchas generaciones de reacciones idiotipo-antiidiotipo. Los humanos sintetizan pequeñas cantidades de acs contra secuencias de proteínas tisulares (auto-acs) y a su turno, estimulan la producción de acs antiidiotipícos contra ellos.
Las preparaciones concentradas de IgGIV, contienen gran cantidad de acs capaces de reaccionar con miles de determinantes antigénicos, debido a la experiencia inmunológica adquirida por los 10.000 donantes, con los cuales se fabrican los productos de IgGIV; en medio de tales acs, habrán muchos de especificidad antiidiotípica (3).

Figura 1. Red idiotipo-antiidiotipo. El ag se une a receptores específicos en las CT (CD4+A.1), que con el concurso de moléculas coestimulatorias y citoquinas (MCE+C), proporcionan señales permisivas a las CB (B1), lo que lleva a su diferenciación en células plasmáticas (CP1) y a la secreción de acs. Los acs producidos por CP1 son reconocidos por otras CT inductoras (CD4+A-2 Id2) con receptores específicos de membrana (TCR: receptor CT) para los ags. Estas CT envían señales a otras CB (B2-Id2), capaces de reconocer los determinantes idiotípicos del primer ac (Id2). Las CB se transforman en células plasmáticas (CP2) que sintetizan un ac anti-idiotípico que reacciona con el ac producido por CP1. Este segundo ac (a1d1), es semejante al ag y puede unirse al receptor para el ag en las CB (B1). Esta interacción puede aumentar la formación de este primer ac pero, más a menudo lo suprime. a1d1 será reconocido por CT y B específicas, lo que induce una reacción en cadena, pero a1d1 se puede unir a receptores (TCR) específicos de CT reguladoras (CD4+R(-)Id). Después de esta interacción, se suprimen las actividades de CT y B, lo que origina un bloqueo de la reacción original frente al ag.
Además de la IgG, las preparaciones de IgGIV contienen otros “factores contaminantes”: moléculas de membrana y receptores solubilizados (CD4, CD5, receptor del complemento CR1), antígenos de histocompatibilidad: HLA clase I y II; acs específicos contra determinantes del HLA y otras moléculas de superficie de los linfocitos (anti CD4, CD8), acs contra citoquinas y sus receptores, acs contra superantígenos.
Estas moléculas son importantes en la regulación de las respuestas inmunes inflamatorias y en las enfermedades autoinmunes (4).
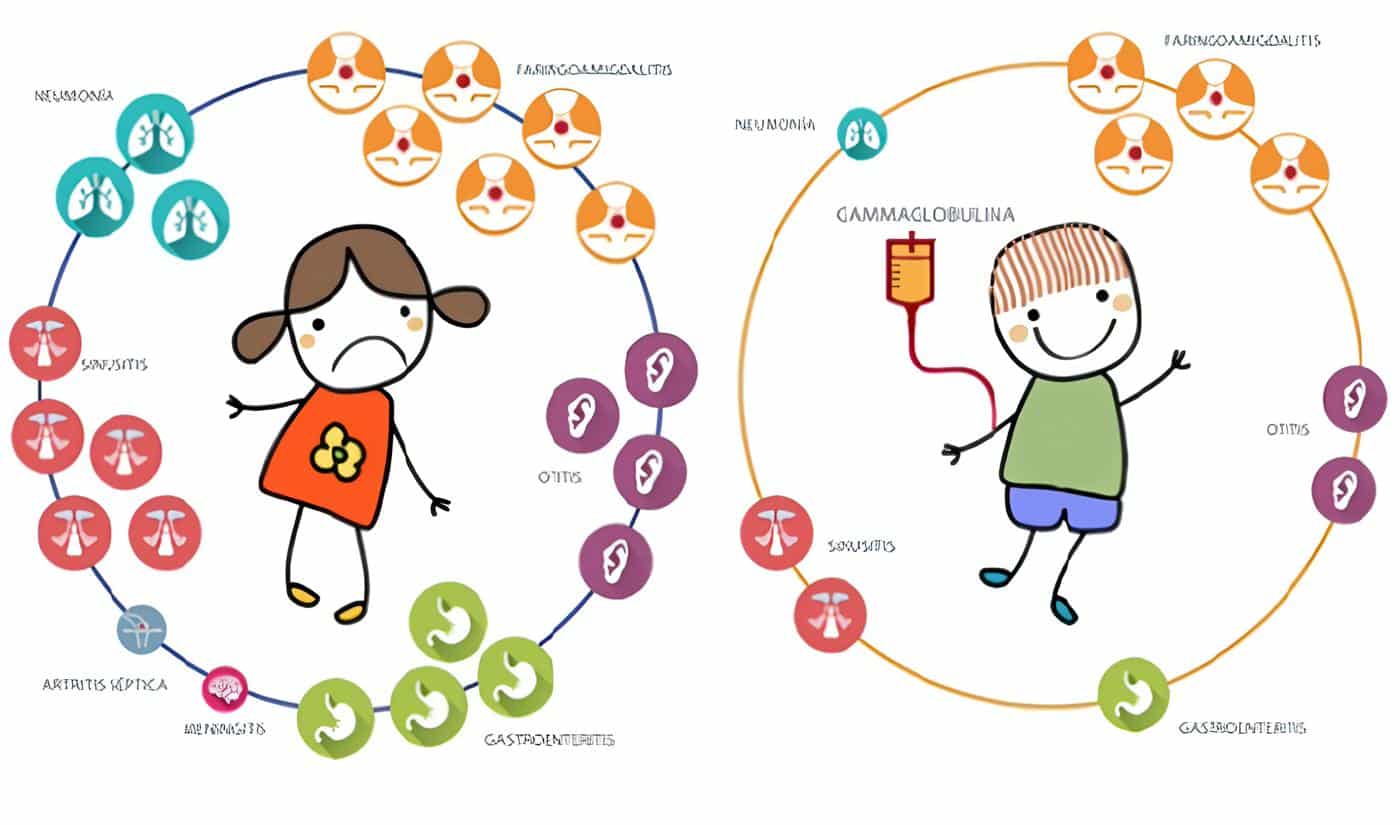
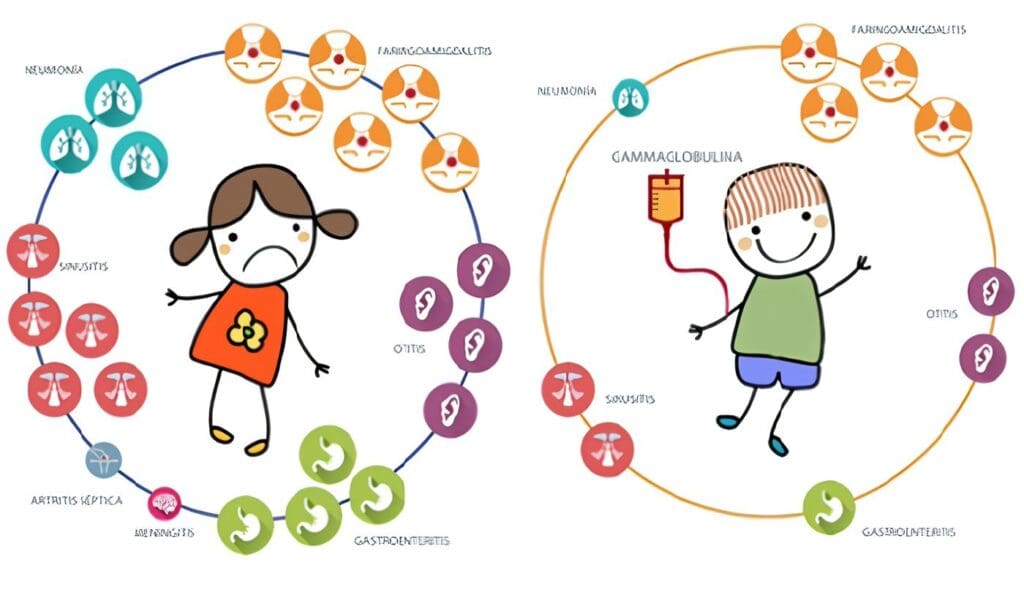
La IgGIV se emplea como terapia de reemplazo en inmunodeficiencias primarias, SIDA en niños, infecciones en el recién nacido de bajo peso, trasplante de médula ósea y leucemia linfocítica crónica (5).
Pero el uso de la IgGIV ha experimentado un gran crecimiento en años recientes; este rápido desarrollo se debe al progreso en la calidad de las presentaciones, lo cual ha reducido la morbilidad por su aplicación, y especialmente al incremento de los informes sobre los beneficios que se han alcanzado, y los que se pueden obtener con la inmunomodulación de la fisiopatología de muchas enfermedades.
De gran interés ha sido el descubrimiento de la modificación drástica de las reacciones inmunes, por la administración de grandes cantidades de IgGIV: 400 a 2000 mg/Kg de peso corporal, en un período de dos a cinco días.
Muchos de los resultados que se han obtenido en la manipulación de la IgGIV sobre el sistema inmune, sugieren que las porciones Fc y el idiotipo de estas moléculas, son las que ejercen dichas acciones (6, 7).
(Lea También: Bloqueo y Modulación de los Receptores FC)
Inmunomodulación por la IgGIV
La IgGIV puede ejercer múltiples acciones reguladoras de la respuesta inmune. Los principales mecanismos establecidos son los siguientes:
1. Bloqueo y modulación de los receptores Fc
- Bloqueo de los receptores Fcg presentes en las células fagocíticas.
- Disminución de la afinidad de los receptores Fcg de las células fagocíticas.
- Incremento de los niveles de CD16 soluble.
2. Modulación de la producción y acción de las citoquinas
- Inhibición de la producción y secreción de citoquinas por macrófagos y CT: IL-1, IL-2, IL-3, IL-4, IL-5, IL-6, IL-10, FEC-GM (Factor estimulante de colonias granulocito y monocito), FNT (Factor de necrosis de tumores), IFNg.
- Aumento de la producción de IL-8 y del antagonista del receptor de IL-1 (IL-1 Ra)
- Acción de acs contra citoquinas (IL-1, FNT a).
- Disminución de la expresión del receptor para IL-2.
3. Inhibición de la función de las células B
- Inhibición de la activación de las CB y disminución de la producción de Igs por medio del receptor FcgRIIb.
- Incremento de la apoptosis de las CB.
4. Modulación de la función de las células T
- Disminución de la respuesta proliferativa de las CT ante el reto con mitógenos y antígenos.
- Inhibición de la activación de las CT mediada por superantígenos.
- Interferencia de la comunicación entre las células presentadoras de antígeno y las CT, por los correceptores solubles (CD5, CD4., CD8), las moléculas del HLA clase I y II, y los anticuerpos anti CD4 y anti HLA.
5. Restauración de la red idiotípica
- Ejerce interacciones reguladoras idiotipo-antiidiotipo restaurando la red idiotípica.
- Acs contra auto-acs: antifactor VIII, antitiroglobulina, antiADN, antifactor intrínseco, ANCAs, y otros.
6. Regulación de la actividad del complemento
- Modulación de la incorporación de los factores derivados del complemento sobre las células blanco (C3b, C4b, complejo de ataque a membrana).
- Posee receptores solubles para el complemento: CR1.
- Favorece la solubilización de complejos inmunes circulantes y depositados en los tejidos (1, 4, 8-15)