Dr. Germán Barón Castañeda, M.D.
El gene para la subunidad a de la gonadotropina es expresado tanto en la hipófisis como en la placenta. La subunidad b de la HCG es expresada en la placenta pero no en la hipófisis y la subunidad b de la LH se encuentra en la hipófisis pero no en la placenta.
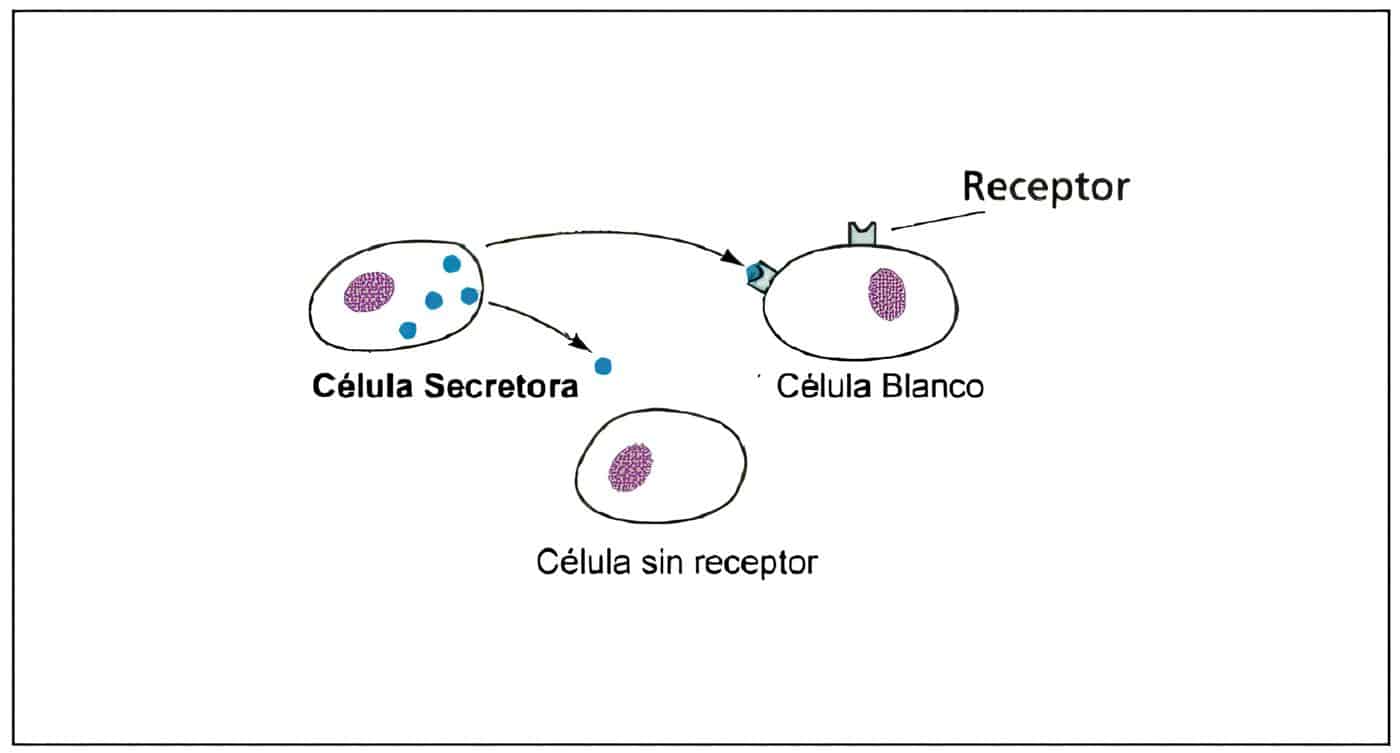
Ambas gonadotropinas son secretadas en la misma célula, el gonadotropo, localizada especialmente en las porciones laterales de la glándula hipófisis. La población de gonadotropos a nivel hipofisiario es heterogénea, teniendo unos producción de FSH, otros de LH y otros de ambas gonadotropinas.
La GnRH actúa sobre receptores específicos en los gonadotropos hipofisiarios. Inicialmente los receptores están distribuidos en forma regular sobre la superficie celular. La unión de la GnRH con su receptor induce la formación de nuevos receptores que luego son “internalizados”.
El complejo hormona-receptor es degradado en el interior de la célula y gran número de receptores regresa a la superficie celular. Este fenómeno es conocido como regulación hacia arriba. Si la exposición a la GnRH es continua, los receptores “internalizados” no retornan a la superficie, produciendo la regulación hacia abajo.
Los receptores de GnRH son regulados por muchas sustancias, entre ellas incluidas la misma GnRH, la inhibina, la activina y los esteroides sexuales.
Los gonadotropos
La acción de la GnRH sobre los gonadotropos está mediada por mecanismos dependientes de calcio/calmodulina.
Se requiere de la acción de un péptido extracelular, la proteína G, que permite el intercambio de GDP a GTP.
La proteína G al unirse al GTP activa la enzima fosfolipasa C, generando la producción de dos segundos mensajeros, el bifosfato de fosfoinositol y el 1,2-diacilglicerol. El primero de ellos aumenta el calcio intracelular y desencadena la liberación de las gonadotropinas preformadas por exocitosis.
El diacilglicerol activa la protein-quinasa C, lo cual induce los genes que regulan la síntesis de gonadotropinas.
La acción de la GnRH sobre sus receptores activa múltiples mensajeros, produciendo inicialmente la liberación de gonadotropinas previamente almacenadas.
Por el fenómeno de “auto-cebamiento” se sintetizan nuevos receptores que permiten que la célula responda a cantidades menores de GnRH, evento importante para la aparición del pico de LH previo a la ovulación; este fenómeno es mayor en la fase folicular avanzada, lo cual permite inferir que es potenciado por los estrógenos. El factor limitante en la síntesis de gonadotropinas es la disponibilidad de subunidades b.
Péptidos cerebrales
Diversas clases de péptidos se encuentran en el sistema nervioso, algunos de ellos diseminados por todo el organismo. Hasta ahora se empiezan a dilucidar las posibles acciones que tienen en la regulación cerebral sobre la hipófisis.
- Neurotensina: Es un vasodilatador. Altera la liberación de hormonas hipofisiarias y disminuye la temperatura corporal.
- Colecistoquinina: puede estar comprometida en la regulación del comportamiento, saciedad e ingesta de líquidos.
- Péptido intestinal vasoactivo (VIP): produce vasodilatación, conversión de glucógeno a glucosa, lipólisis y secreción de insulina. En la hipófisis es sintetizado en los lactotropos y aumenta la secreción de prolactina.
- Angiotensina II: Receptores para ella se encuentran en varios tipos de células hipofisiarias. A nivel hipotalámico parece influir en los efectos de la noradrenalina y la dopamina sobre las hormonas liberadoras.
- Endotelina: Puede inducir la liberación de vasopresina de la hipófisis posterior y de gonadotropinas de la anterior. Inhibe la respuesta de prolactina.
-
Somatostatina: Inhibe la liberación de hormona de crecimiento, prolactina, ACTH y TSH en la hipófisis. Está localizada en neuronas sensoriales y puede actuar como transmisor del dolor.
- Neuropéptido Y: Su secreción en el hipotálamo es regulada por esteroides sexuales. Estimula la liberación pulsátil de GnRH y potencia la respuesta hipofisiaria a GnRH. En ausencia de estrógenos bloquea la secreción de hormona liberadora.
- Factores de crecimiento: Modulan la producción y secreción de hormonas hipofisiarias.
- Activina e inhibina: La inhibina inhibe de forma selectiva la secreción de FSH pero no de LH. La activina aumenta la secreción de FSH e inhibe la prolactina y la hormona de crecimiento.
- Folistatina: Es secretada por una variedad de células hipofisiarias, incluyendo los gonadotropos. Su principal acción es inhibir la síntesis y secreción de FSH, además de su respuesta a GnRH.
- Galanina: Es producida en los lactotropos y liberada a la circulación portal en forma pulsátil. Estimula la secreción de LH. Su secreción es inhibida por la dopamina y somatostatina y estimulada por TRH y estrógenos.
- Opioides endógenos: La producción de opioides está regulada por la síntesis de péptidos precursores, de los cuales existen 3: Proopiomelanocortina, fuente de endorfinas, Proencefalina A y B, fuente de encefalinas y Prodinorfina, fuente de las dinorfinas.
(Lea También: Sistema TRH/TSH, Sistema Neurohipofisiario y Glándula Pineal)
De la proopiomelanocortina se derivan el precursor de ACTH y la b-lipotropina; esta última se rompe en varios fragmentos, hormona estimulante de los melanocitos, encefalina y endorfinas a, b y g. De estas, la que se encuentra en mayor cantidad en el hipotálamo es la b-endorfina.
La b-endorfina regula varias funciones hipotalámicas incluyendo la reproducción, temperatura, función cardiovascular y respiratoria y otras centrales tales como la percepción del dolor y el ánimo.
La expresión del gene de la proopiomelanocortina a nivel hipofisiario es controlada por la hormona liberadora de corticotropina (CRH) y afectada por la retroalimentación negativa de glucocorticoides. En el hipotálamo es regulada por la presencia de esteroides sexuales.
Los opioides tienen variación a lo largo del ciclo menstrual, siendo sus niveles más bajos en la época menstrual y aumentando progresivamente hasta la fase lútea. Una reducción en la frecuencia de los pulsos de LH está asociada con aumento en la secreción de endorfina.
Por lo tanto, los opioides endógenos inhiben la secreción de gonadotropinas al suprimir la liberación hipotalámica de GnRH. Los esteroides sexuales actúan por retroalimentación sobre los opioides, frenando de esta manera la liberación de gonadotropinas.
Tanto los estrógenos como la progesterona en forma aislada aumentan los opioides endógenos, pero los estrógenos facilitan la acción de la progesterona, lo cual explica la máxima supresión de GnRH durante la fase lútea.
La acción de la encefalina está relacionada con la modulación de la vía de las catecolaminas, especialmente de norepinefrina. No intervienen receptores para dopamina, acetilcolina o a-adrenérgicos. Posiblemente la endorfina puede afectar directamente la liberación de GnRH.
Catecolestrógenos
Son derivados de los estrógenos gracias a la acción de la enzima 2-hidroxilasa. Tienen dos caras, una catecol y otra estrógeno, por lo cual tienen la capacidad de actuar con receptores para ambas sustancias. Posiblemente afectan la secreción de GnRH, función que aún es especulativa.
 Figura 2.1 Control hipotálamo-hipofisiario
Figura 2.1 Control hipotálamo-hipofisiario
Tanicitos
Son células ependimarias especializadas cuyos cuerpos ciliados tapizan el tercer ventrículo. Pueden transportar sustancias desde el líquido céfaloraquideo al sistema porta hipofisiario. Los tanicitos cambian de morfología en respuesta a esteroides y durante el ciclo ovárico.
El sistema de CRH – ACTH
Actualmente se conoce la estructura bioquímica de la Hormona liberadora de corticotropina (CRH), compuesta por 41 aminoácidos y codificada por un gen localizado en el brazo largo del cromosoma 8.
La vía de la CRH que controla la secreción hipofisiaria de ACTH se encuentra en el núcleo paraventricular y termina en la eminencia media. Este núcleo se encuentra densamente inervado por fibras adrenérgicas y noradrenérgicas medulares y contiene receptores adrenérgicos. Existen también neuronas en el área preóptica del hipotálamo que pueden actuar como neurotransmisores excitadores. Se ha encontrado esta hormona en la médula suprarrenal y la placenta.
La liberación de CRH y por ende de ACTH está regulada por los niveles de cortisol plasmático. En condiciones de estrés llegan estímulos adrenérgicos al núcleo paraventricular a través de vías ascendentes del tronco del encéfalo. La interleuquina-1 puede actuar como intermediario entre el sistema inmune y la liberación de CRH.
Otras vías que intervienen en el control de la CRH incluyen:
- La proopiomelanocortina: los opioides suprimen la liberación de CRH.
- La vía angiotensinérgica
- La vía del neuropéptido Y que actúa como estimuladora independiente de las interacciones catecolaminérgicas.
La acción de la CRH en la hipófisis implica la unión a receptores específicos que utilizan el AMPc como segundo mensajero. Se ha demostrado la presencia de receptores en el área preóptica y en el núcleo periventricular del hipotálamo; de esta manera puede llegar a las terminaciones de la GnRH y hormona de crecimiento y afectar su secreción.
La secreción de CRH tiene importancia en la reproducción, ya pues en situaciones de estrés puede bloquear la liberación de GnRH.
Sistema GHRH – Hormona de crecimiento
La secreción de hormona de crecimiento se encuentra bajo la influencia de múltiples estímulos externos como el ejercicio, el estrés, las comidas ricas en proteínas, la hipoglicemia y el sueño. Su producción se caracteriza por la liberación episódica; aumenta en frecuencia y amplitud durante la pubertad. Su secreción es estimulada por los estrógenos, testosterona y hormona tiroidea e inhibida por los niveles elevados de glucocorticoides.
El control hipotalámico es ejercido a través de dos péptidos: la somatostatina y el factor liberador de hormona de crecimiento (GHRH). Las neuronas productores de GHRH se encuentran localizadas principalmente en el núcleo periventricular posterior y sus terminaciones se proyectan hacia la eminencia media.
A nivel periférico juega papel importante en la regulación del factor de crecimiento similar a la insulina I.
La vía de la somatostatina que controla la hormona de crecimiento se encuentra en los núcleos periventriculares y paraventriculares. Su secreción ejerce una potente inhibición sobre la producción de hormona de crecimiento.
Tiene algún efecto fisiológico bloqueando la liberación de TSH. La somatostatina no solo está en el sistema nervioso; se encuentra también en el tracto gastrointestinal, el páncreas y la placenta. Actúa sobre el flujo sanguíneo y la motilidad intestinal y posiblemente tiene un efecto supresor sobre el sistema inmune.
El mecanismo de acción de la GHRH en la hipófisis se ejerce a través del AMPc. La somatostatina actúa por medio de una proteína G inhibitoria que bloquea los canales del calcio y la subunidad catalítica de la adenil-ciclasa. La respuesta hipofisiaria a la GHRH depende de la edad, siendo casi nula en mayores de 40 años.