La granulomatosis de Wegener es una vasculitis sistémica, que generalmente afecta el tracto respiratorio alto y bajo y el riñón, la cual se caracteriza por la presencia de anticuerpos anti-neutrófilos. Las interacciones entre estos anticuerpos y los FcgR en los monocitos se ha asociado con la inducción de factores quimiotácticos para neutrófilos, tales como la IL-8. La activación de los neutrófilos vía los FcgRIIIB con la consecuente activación de la explosión respiratoria, inician y promueven el proceso inflamatorio en los vasos sanguíneos que es responsable de los cambios histopatológicos de esta enfermedad (49).
La TIH es una forma de trombocitopenia observada ocasionalmente después del tratamiento con heparina y se debe a la FORMACIÓN de anticuerpos contra los complejos Heparina/factor plaquetario 4 (Heparina/FP4) (50). La heparina tiene una estructura a base de polisacáridos que le permite inducir la formación de anticuerpos tipo IgG2.
El FcgRIIA es expresado por las plaquetas y la interacción de estos receptores con los complejos inmunes forman agregados más grandes que son eliminados por el sistema de fagocitos mononucleares y de esta forma se depleta las reservas de plaquetas.
La TIH y los eventos trombóticos se asocian con el fenotipo FcgRIIAR/R131 pues la disminución en la depuración de los complejos Heparina/FP4 en los pacientes portadores de este alelo permite la prolongación de la activación de las plaquetas y del endotelio, aumentando el riesgo de complicaciones trombóticas (50, 51).
FcgR y alergia
Es claro que en la hipersensibilidad tipo I, el receptor que juega un papel protagónico es el FceRI, sin embargo, los FcgR también tienen un papel importante en la génesis de dichas reacciones.
Por ejemplo, los ratones deficientes de FcgRIIB presentan un incremento de las respuestas anafilácticas mediadas por IgE, lo que sugiere una relación entre este receptor inhibitorio y la vía del FceRI (52). Los mastocitos, además de expresar en su membrana FceRI, también expresan FcgRIIB y FcgRIII y hay evidencias que indican que la IgE puede unirse con baja afinidad a FcgRII y al FcgRIII (52).
Además la IgG tiene la capacidad de activar los mastocios por medio de FcgRIII. ESTUDIOS indican que la cadena g es muy importante regulando el nivel de expresión de FceRI y FcgRIII, pues parece ser la competencia por las distintas cadenas la que determina la expresión en membrana de cada receptor y su habilidad para responder a los distintos estímulos biológicos.
El nivel absoluto de expresión de los FcRs en el mastocito tiene en consecuencia una importancia terapéutica amplia, ya que tanto la IgG como la IgE modulan la respuesta inflamatoria, por eso la modulación de la expresión de la cadena g podría ser alternativa terapéutica en enfermedades como anafilaxis y asma.
La Inmunoglobulina Intravenosa como Modulador de la Expresión de FcgR
Las preparaciones de inmunoglobulinas para uso intravenoso (IgIV) han sido utilizadas como moduladores del curso de enfermedades autoinmunes-inflamatorias, antitoxinas y antimicrobinos (53).
Se ha comprobado que las porciones monoméricas de las fracciones Fc, pero no la porción Fab de la IgG, son las que confieren un efecto protector en modelos de artritis y púrpura trombocitopénico autoinmune (54).
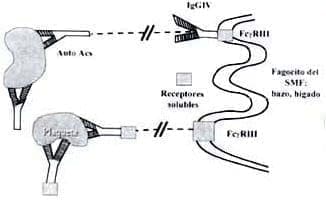
Estos hallazgos han permitido asociar los efectos de la aplicación de la IgIV con los receptores Fc. La PROTECCIÓN o mejoría de la respuesta inflamatoria que se observa en estos modelos se ha asociado con la capacidad que tiene la IgIV de inducir la expresión de FcgRIIB en los macrófagos (54).
Las evidencias sugieren que ocurre un aumento del 60% en el número de macrófagos que expresan este receptor inhibitorio, pero sin que hubiera un aumento de la cantidad de receptores expresados por estas células (53). La modulación de este receptor aparece como posible diana terapéutica en las enfermedades inflamatorias desencadenadas por anticuerpos.
Conclusión
La información que se tiene sobre los receptores para la porción Fc de la IgG es amplia y cada vez se aclara más su fisiología y estructura.
Se entiende como estos receptores por tener múltiples funciones también pueden estar involucrados en la génesis de varias alteraciones.
Sin embargo, dado el gran dinamismo y la superposición de funciones durante la respuesta inmune, es difícil que una alteración en estos receptores explique por sí misma todo un cuadro clínico.
Estos receptores no deben ser entendidos de manera aislada; simultáneamente a la activación de los receptores Fcg se están induciendo e inhibiendo la señalización desde otros receptores en la membrana y en el citosol, de forma tal que la respuesta biológica depende de múltiples variables.
Por ello es necesario el ESTUDIO de otros receptores y proteínas del sistema inmune que eventualmente puedan presentar polimorfismos desventajosos para el individuo que las porta.
Esto podría ayudar a entender porque individuos con una misma enfermedad presentan cuadros clínicos que varían en presentación y en intensidad. Además, el conocimiento acerca de la relación antígeno-anticuerpo-FcgR y la señalización intracelular que se desencadena tras dicha interacción es de gran importancia y se perfila como blanco de intervención terapéutica futura.
Referencias Bibliográficas
- 1. van der Pol W, van de Winkel JG. IgG receptor polymorphisms: risk factors for disease. Immunogenetics 1998;48(3):222-32.
- 2. Fridman WH, Teillaud JL, Bouchard C, Teillaud C, Astier A, Tartour E, et al. Soluble Fc gamma receptors. J Leukoc Biol 1993;54(5):504-12.
- 3. Mathur A, Lynch RG, Kohler G. Expression, distribution and specificity of Fc receptors for IgM on murine B cells. J Immunol 1988;141(6):1855-62.
- 4. Letourneur F, Hennecke S, Demolliere C, Cosson P. Steric masking of a dilysine endoplasmic reticulum retention motif during assembly of the human high affinity receptor for immunoglobulin E. J Cell Biol 1995;129(4):971-8.
- 5. Lobell RB, Arm JP, Raizman MB, Austen KF, Katz HR. Intracellular degradation of Fc gamma RIII in mouse bone marrow culture-derived progenitor mast cells prevents its surface expression and associated function. J Biol Chem 1993;268(2):1207-12.
- 6. Kurosaki T, Gander I, Wirthmueller U, Ravetch JV. The beta subunit of the Fc epsilon RI is associated with the Fc gamma RIII on mast cells. J Exp Med 1992;175(2):447-51.
- 7. Masuda M, Roos D. Association of all three types of Fc gamma R (CD64, CD32, and CD16) with a gamma-chain homodimer in cultured human monocytes. J Immunol 1993;151(12):7188-95.
- 8. van de Winkel JG, Ernst LK, Anderson CL, Chiu IM. Gene organization of the human high affinity receptor for IgG, Fc gamma RI (CD64). Characterization and evidence for a second gene. J Biol Chem 1991;266(20):13449-55.
- 9. Osman N, Kozak CA, McKenzie IF, Hogarth PM. Structure and mapping of the gene encoding mouse high affinity Fc gamma RI and chromosomal location of the human Fc gamma RI gene. J Immunol 1992;148(5):1570-5.
- 10. van de Winkel JG, Capel PJ. Human IgG Fc receptor heterogeneity: molecular aspects and clinical implications. Immunol Today 1993;14(5):215-21.
- 11. Hulett MD, Osman N, McKenzie IF, Hogarth PM. Chimeric Fc receptors identify functional domains of the murine high affinity receptor for IgG. J Immunol 1991;147(6):1863-8.
- 12. van Vugt MJ, Heijnen AF, Capel PJ, Park SY, Ra C, Saito T, et al. FcR gamma-chain is essential for both surface expression and function of human Fc gamma RI (CD64) in vivo. Blood 1996;87(9):3593-9.
- 13. Miller KL, Duchemin AM, Anderson CL. A novel role for the Fc receptor gamma subunit: enhancement of Fc gamma R ligand affinity. J Exp Med 1996;183(5):2227-33.
- 14. van Vugt MJ, Kleijmeer MJ, Keler T, Zeelenberg I, van Dijk MA, Leusen JH, et al. The FcgammaRIa (CD64) ligand binding chain triggers major histocompatibility complex class II antigen presentation independently of its associated FcR gamma-chain. Blood 1999;94(2):808-17.
- 15. Ceuppens JL, Baroja ML, Van Vaeck F, Anderson CL. Defect in the membrane expression of high affinity 72-kD Fc gamma receptors on phagocytic cells in four healthy subjects. J Clin Invest 1988;82(2):571-8.
- 16. van de Winkel JG, de Wit TP, Ernst LK, Capel PJ, Ceuppens JL. Molecular basis for a familial defect in phagocyte expression of IgG receptor I (CD64). J Immunol 1995;154(6):2896-903.
- 17. Qiu WQ, de Bruin D, Brownstein BH, Pearse R, Ravetch JV. Organization of the human and mouse low-affinity Fc gamma R genes: duplication and recombination. Science 1990;248(4956):732-5.
- 18. Warmerdam PA, Nabben NM, van de Graaf SA, van de Winkel JG, Capel PJ. The human low affinity immunoglobulin G Fc receptor IIC gene is a result of an unequal crossover event. J Biol Chem 1993;268(10):7346-9.
- 19. Metes D, Ernst LK, Chambers WH, Sulica A, Herberman RB, Morel PA. Expression of functional CD32 molecules on human NK cells is determined by an allelic polymorphism of the FcgammaRIIC gene. Blood 1998;91(7):2369-80.
- 20. Hulett MD, Witort E, Brinkworth RI, McKenzie IF, Hogarth PM. Multiple regions of human Fc gamma RII (CD32) contribute to the binding of IgG. J Biol Chem 1995;270(36):21188-94.
- 21. Brooks DG, Qiu WQ, Luster AD, Ravetch JV. Structure and expression of human IgG FcRII(CD32). Functional heterogeneity is encoded by the alternatively spliced products of multiple genes. J Exp Med 1989;170(4):1369-85.
- 22. Warmerdam PA, van de Winkel JG, Gosselin EJ, Capel PJ. Molecular basis for a polymorphism of human Fc gamma receptor II (CD32). J Exp Med 1990;172(1):19-25.
- 23. Stuart SG, Simister NE, Clarkson SB, Kacinski BM, Shapiro M, Mellman I. Human IgG Fc receptor (hFcRII; CD32) exists as multiple isoforms in macrophages, lymphocytes and IgG-transporting placental epithelium. Embo J 1989;8(12):3657-66.
- 24. Muta T, Kurosaki T, Misulovin Z, Sanchez M, Nussenzweig MC, Ravetch JV. A 13-amino-acid motif in the cytoplasmic domain of Fc gamma RIIB modulates B-cell receptor signalling. Nature 1994;368(6466):70-3.
- 25. Van den Herik-Oudijk IE, Ter Bekke MW, Tempelman MJ, Capel PJ, Van de Winkel JG. Functional differences between two Fc receptor ITAM signaling motifs. Blood 1995;86(9):3302-7.
- 26. Van den Herik-Oudijk IE, Capel PJ, van der Bruggen T, Van de Winkel JG. Identification of signaling motifs within human Fc gamma RIIa and Fc gamma RIIb isoforms. Blood 1995;85(8):2202-11.
- 27. Van Den Herik-Oudijk IE, Westerdaal NA, Henriquez NV, Capel PJ, Van De Winkel JG. Functional analysis of human Fc gamma RII (CD32) isoforms expressed in B lymphocytes. J Immunol 1994;152(2):574-85.
- 28. Warmerdam PA, van de Winkel JG, Vlug A, Westerdaal NA, Capel PJ. A single amino acid in the second Ig-like domain of the human Fc gamma receptor II is critical for human IgG2 binding. J Immunol 1991;147(4):1338-43.