Las hemoglobinas son proteínas globulares, presentes en los hematíes en altas concentraciones, que fijan oxígeno en los pulmones y lo transportan por la sangre hacia los tejidos y células que rodean el lecho capilar del sistema vascular. Al volver a los pulmones, desde la red de capilares, la hemoglobina actúa como transportador de CO2 y de protones. Este es uno de los ejemplos más llamativos de la relevancia del proceso evolutivo y la eficiencia de los sistemas biológicos.
Los diferentes tipos de cadenas de la hemoglobina se denominan alfa, beta, gamma y delta. La hemoglobina A –la forma más frecuente en el adulto- es una combinación de dos cadenas alfa y dos cadenas beta. Debido a que cada cadena tiene un grupo proteico hem, hay 4 átomos de hierro en cada molécula de hemoglobina; cada una de ellas puede unirse a una molécula de oxígeno, siendo pues un total de 4 moléculas de oxígeno las que pueden transportar cada molécula de hemoglobina.
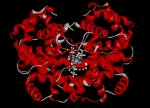 Las cuatro cadenas polipeptídicas de la Hb contienen cada una un grupo prostético hem. Un grupo prostético es la porción no polipeptídica de una proteína. El hem es una molécula de porfirina que contiene un átomo de hierro en su centro. El tipo de porfirina de la Hb es la protoporfirina IX; contiene dos grupos ácidos propiónicos, dos vinilos y cuatro metilos como cadenas laterales unidas a los anillos pirrólicos de la estructura de la porfirina. El átomo de hierro se encuentra en estado de oxidación ferroso (+2) y puede formar cinco o seis enlaces de coordinación dependiendo de la unión del O2 (u otro ligando) a la Hb (oxiHb, desoxiHb). Cuatro de estos enlaces se producen con los nitrógenos pirrólicos de la porfirina en un plano horizontal. El quinto enlace de coordinación se realiza con el nitrógeno del imidazol de una histidina denominada histidina proximal. Finalmente, el sexto enlace del átomo ferroso es con el O2, que además está unido a un segundo imidazol de una histidina denominada histidina distal. Tanto el quinto como el sexto enlace se encuentran en un plano perpendicular al plano del anillo de porfirina. Las cadenas polipeptídicas alfa contienen 141 aminoácidos, las no a 146 (b, g, d) y difieren en la secuencia de aminoácidos. Se conoce desde hace décadas la estructura primaria de las cuatro cadenas de Hb normales.
Las cuatro cadenas polipeptídicas de la Hb contienen cada una un grupo prostético hem. Un grupo prostético es la porción no polipeptídica de una proteína. El hem es una molécula de porfirina que contiene un átomo de hierro en su centro. El tipo de porfirina de la Hb es la protoporfirina IX; contiene dos grupos ácidos propiónicos, dos vinilos y cuatro metilos como cadenas laterales unidas a los anillos pirrólicos de la estructura de la porfirina. El átomo de hierro se encuentra en estado de oxidación ferroso (+2) y puede formar cinco o seis enlaces de coordinación dependiendo de la unión del O2 (u otro ligando) a la Hb (oxiHb, desoxiHb). Cuatro de estos enlaces se producen con los nitrógenos pirrólicos de la porfirina en un plano horizontal. El quinto enlace de coordinación se realiza con el nitrógeno del imidazol de una histidina denominada histidina proximal. Finalmente, el sexto enlace del átomo ferroso es con el O2, que además está unido a un segundo imidazol de una histidina denominada histidina distal. Tanto el quinto como el sexto enlace se encuentran en un plano perpendicular al plano del anillo de porfirina. Las cadenas polipeptídicas alfa contienen 141 aminoácidos, las no a 146 (b, g, d) y difieren en la secuencia de aminoácidos. Se conoce desde hace décadas la estructura primaria de las cuatro cadenas de Hb normales.
Para operar como vehículo de intercambio gaseoso, la hemoglobina (Hb) debe satisfacer ciertos requerimientos básicos como son: ser capaz de transportar cantidades considerables de oxígeno; ser muy soluble; captar y descartar oxígeno a presiones apropiadas y, ser un buen amortiguador. Más del 95% de la hemoglobina del adulto y de los niños mayores de 7 meses es A (HbA). Su estructura se designa como a2b2, para indicar que posee dos cadenas a y dos b. Los adultos normales también tienen un 2-3% de HbA2, la cual está compuesta por dos cadenas a como las de la Hb A, y dos cadenas d. Se representa como a2d2. Las cadenas d son diferentes de las cadenas b y están bajo control genético independiente. Normalmente también existen diversas especies de HbA modificada; se designan como A1a1, A1a2, A1b y A1c, y se deben a modificaciones postraduccionales de la Hb con diversos azúcares como glucosa-6-fosfato. La más importante cuantitativamente es la Hb A1c. Proviene de la fijación covalente de un resto de la glucosa al extremo N-terminal de la cadena b. La reacción no es catalizada enzimáticamente, dependiendo entonces su velocidad de la concentración de glucosa. Por tanto, la Hb A1c constituye una medida útil del control de los pacientes diabéticos durante los días o semanas previos a la toma de la muestra. La Hb fetal (HbF), es el componente principal de la Hb del recién nacido. Posee dos cadenas g en lugar de dos cadenas b y se representa a2g2. La HbF está adaptada al ambiente materno-fetal. Ha de fijar el oxígeno mucho más fuerte para competir por el O2 con la HbA materna (su curva de saturación está desplazada a la izquierda). Esto se consigue gracias a que dos de los grupos que recubren la cavidad de fijación de 2,3-difosfoglicerato (DPG) tienen cadenas laterales neutras, a diferencia de la HbA en la que están cargadas positivamente. Lo anterior hace que el DPG, cargado negativamente, se una menos a la HbF y en lugar de ello, se fije más fuerte el O2. Además, entre 15% y 20% de la HbF está acetilada en sus N-terminales y se denomina HbF1; esta variante no fija DPG.
Las hemoglobinas están presentes en todos los reinos de la naturaleza con particulares especializaciones de acuerdo con las necesidades de cada organismo. Se pueden sintetizar grandes cantidades de hemoglobina en células especializadas, como los eritrocitos, cuando es necesario transportar eficientemente O2 a sitios especiales de intensa actividad respiratoria. También se generan cantidades pequeñas de proteínas como la mioglobina y la hemoglobina no simbiótica de las plantas cuando es necesario un rápido sistema intracelular de liberación de O2, como en la mitocondria o el cloroplasto. Cuando el sistema de transporte de electrones no está separado del compartimiento intracelular, como en las bacterias, las hemoglobinas pueden ser usadas, bajo condiciones hipóxicas, para liberar oxígeno como aceptor terminal de electrones. Por tanto, existe una conexión evolutiva entre las hemoglobinas que transportan O2, los citocromos que pasan electrones a través de la cadena respiratoria y la fotosíntesis, y las demás hemoproteínas que catalizan reacciones redox. Quizá 1.800 millones de años atrás los anillos de porfirina unidos a un metal evolucionaron junto con la fotosíntesis y la consiguiente aparición del O2 en la atmósfera, a partir de un gen ancestral común.
Pronto en la evolución se incorporó la utilidad de los anillos de porfirina unidos a un metal para la transferencia de electrones. Tal es el caso de los citocromos que transportan electrones en la cadena respiratoria, u otras hemoproteínas que catalizan una clase variada de reacciones de óxido-reducción. Aun en la fotosíntesis, los anillos de porfirina transportan los electrones generados a partir de la luz solar y el agua para luego generar ATP. Otras hemoproteínas se han adaptado para unir reversiblemente el oxígeno generado en la fotosíntesis y transportarlo a cada célula de todos los grupos de organismos: procariotas, hongos, plantas y animales.
La síntesis de hemoglobina comienza en los proeritroblastos y continúa levemente, incluso en el estadio de reticulocito, porque cuando éstos dejan la médula ósea y pasan al torrente sanguíneo, continúan formando cantidades mínimas de hemoglobina durante un día aproximadamente. En primer lugar, la succinil-CoA, formada en el ciclo de Krebs, se une a la glicina para formar una molécula de pirrol. Después, cuatro pirroles se combinan para formar la protoporfirina IX, que tiende a combinarse con el hierro para formar la molécula hem. Finalmente, cada molécula hem se combina con una larga cadena polipéptica, llamada globina, sintetizada por los ribosomas, formando una subunidad llamada cadena de hemoglobina. Cada una de estas cadenas tiene un peso molecular de aproximadamente de 16000; se unen cuatro de ellas de forma laxa para formar la molécula completa de hemoglobina.
Peñuela OA. Hemoglobina: una molécula modelo para el investigador. Colombia Médica 2005; 36: 215-225