Non-syndromic deafness
Leonardo E. Ordóñez, MD*; Alejandra Correa, MD**; Jorge E. Almario, MD***;
José G. Lora, MD****; José A. Prieto MD*****
* Residente de primer año Servicio Integrado de Otorrinolaringología Hospital Militar Central-Hospital Universitario Clínica San Rafael. Universidad Militar Nueva Granada. Bogotá, D.C.
** Residente de tercer año Servicio Integrado de Otorrinolarin-gología Hospital Militar Central-Hospital Universitario Clínica San Rafael. Universidad Militar Nueva Granada. Bogotá, D.C.
*** Jefe Servicio Otorrinolaringología Hospital Militar Central. Profesor asociado Universidad Militar Nueva Granada. Facultad de Medicina Especialista Otorrinolaringología Hospital Universitario Clínica San Rafael. Bogotá, D.C.
**** Profesor auxiliar Universidad Militar Nueva Granada. Facultad de Medicina. Especialista en entrenamiento en Neurotología Hospital Militar Central. Especialista Otorrinolaringología Hospital Universitario Clínica San Rafael. Bogotá, D.C.
***** Jefe Servicio Otorrinolaringología Hospital Universitario Clínica San Rafael. Profesor asociado Universidad Militar Nueva Granada. Facultad de Medicina. Especialista en Entrenamiento en Neurotología Hospital Militar Central. Bogotá, D.C.
Resumen
La pérdida auditiva es la alteración sensorial más común en humanos, 1 de 1000 nacidos vivos presentan una pérdida severa que al no recibir tratamiento conducirá a alteraciones en la adquisición del lenguaje. La mitad de los casos de sordera congénita se atribuye a factores genéticos. De éstos un 70% se clasifican como no sindromáticos y la forma de transmisión autosómica recesiva es la más frecuente.
El descubrimiento de diferentes mutaciones que conducen a pérdida auditiva a llevado a su vez a aclarar las bases moleculares de la fisiología coclear.
Un mecanismo fundamental en el normal funcionamiento del órgano de Corti es el reciclaje del potasio, ión que entra en la célula ciliada e induce su despolarización, saliendo por la membrana lateral y viajando a través de las células del órgano hasta la estría vascular, lugar en donde nuevamente es secretado hacia la endolinfa. Algunas alteraciones en este mecanismo son las causas más frecuentes de sordera de origen genético, destacándose la mutación en el gen de la conexina 26, una proteína de las uniones brecha (gap junction), responsable de más de la mitad de casos de sordera no sindromática, según reportes en diferentes países.
Alteraciones en moléculas de la membrana celular, citoesqueleto celular, matriz extracelular, factores de transcripción, productos de los genes mitocondriales; se han reportado en casos menos frecuentes de sordera no sindromática.
Palabras clave: sordera, sordera congénita, sordera no sindromática, sordera neurosensorial, conexina 26.
Generalidades
La pérdida auditiva es la alteración sensorial más común en humanos, se estima que más de 70 millones de personas alrededor del mundo tienen una pérdida que afecta la comunicación normal (1). La incidencia de hipoacusia congénita severa es de al menos 1 en 1000 nacidos vivos (1, 2). Aproximadamente la mitad se atribuyen a factores genéticos, clasificándose éstos en, aproximadamente, 70% no sindromático y un 30 % sindromático (3). Para Colombia no se conocen datos epidemiológicos al respecto, sin embargo, en un país vecino, Brasil, un estudio reportó que la hipoacusia no sindromática debido a mutación en un gen específico, Conexina 26, tiene un comportamiento similar al descrito en países desarrollados (4). Se presentan estimaciones basadas en datos demográficos de Colombia para el año 2002 (Tabla 1) (5).
TABLA 1 CASOS ESTIMADOS DE HIPOACUSIA EN NIÑOS COLOMBIANOS AL NACER
| Referencia | Nacimientos esperados 20021 | Niños con hipoacusia severa al nacer2 | Niños con hipoacusia de origen genérico3 | Niños con hipoacusia no sindromática4 |
| Bogotá Colombia |
156476 1166849 |
156 1167 |
78 584 |
55 408 |
La pérdida auditiva no sindromática ocurre aisladamente, mientras que en los casos sindromáticos se asocia con anormalidades en otros sistemas. Se han descrito varios cientos de síndromes que cursan con hipoacusia encontrando alteraciones genéticas en algunas formas frecuentes (1-4, 6).
La hipoacusia no sindromática se puede clasificar según su modo de transmisión en: autosómica dominante, autosómica recesiva, ligado a X y mitocondriales (1-3, 6). Se presenta una clasificación según la etiología de la hipoacusia y se muestran las frecuencias aproximadas según diferentes fuentes (1-3)
En los últimos años se han hecho importantes avances en el diag-nóstico de las hipoacusias no sindromáticas, se han encontrado los locus genéticos, y sus productos proteicos, responsables de un importante número casos. Al día de hoy se han identificado 51 locus para las formas autosómicas dominantes, 17 de los cuales se han clonado; 39 para las autosómicas recesivas, con 17 genes clonados; 8 para las ligadas a X con un gen clonado y 2 para las formas de transmisión mitocondrial que están clonados (6). La nomenclatura internacional utilizada para denominar a los locus genéticos de estas diferentes formas de hipoacusia designa DFNA (Deafness A) a las formas de transmisión autosómica dominante, DFNB (Deafness B) a las autosómicas recesivas y DFN (Deafness) aquellas con trans-misión ligada a X. Adicionalmente se coloca un número consecutivo, según el orden cronológico de su descripción, por ejemplo los locus relacionados con la forma de transmisión autosómica dominante se designan de DFNA1 hasta DFNA51 (6).
Fisiología auditiva
El estudio genético de los pacientes con hipoa-cusia ha ayudado a aclarar y ampliar el conocimiento de la fisiología del oído interno en su porción coclear, incluso se han descrito procesos no sospechados hasta encontrar su alteración (2, 7, 8). La cóclea es el sitio en donde la energía mecánica de los ondas sonoras es convertida en potenciales de acción del nervio coclear, iniciando así la transmisión de la información auditiva hacia los centros del tronco cerebral y a centros superiores en la corteza cerebral, proceso necesario para la comprensión e interpretación de los sonidos. La cóclea está localizada en la porción petrosa del hueso temporal, en su longitud está divida en tres compartimentos: la escala vestibular, la escala media y la escala timpánica. La escala media, contiene endolinfa con altas concentraciones de potasio y bajas concentraciones de sodio; esta discrepancia en la composición electrolítica de los líquidos del oído interno genera una diferencia de potencial eléctrico entre el interior y el exterior de la célula, que juega un papel central en el proceso de transducción de la información que se lleva a cabo en la cóclea. (Figura 1).

Figura 1. Distribución coclear de algunos productos genéticos involucrados en sordera no sindromática.
Los receptores auditivos son las células ciliadas del órgano de Corti. El extremo de las estereocilias de las células ciliadas externas están embebidas en la membrana tectoria,
estas cilias tienen un esqueleto de actina y formas no convencionales de miosina que están fijas a una lámina cuticular rica en actina que a su vez sujeta la estereocilia al citoesqueleto celular. Las estereocilias están ancladas unas a otras cerca de su ápice de forma tal que se mueven en conjunto. Las células ciliadas internas son los receptores primarios y reciben la mayoría de las fibras aferentes del nervio coclear; las células ciliadas externas, reciben la mayor parte de la información eferente del mismo nervio, tienen por función promover la discriminación de frecuencia y amplificación de la señal, de forma que modulan el funcionamiento del receptor primario (2, 7, 8). (Figuras 2 y 3).
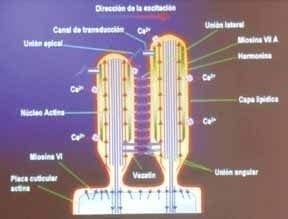 Factura 2. Estructura bio-molecular de la estereocilia.
Factura 2. Estructura bio-molecular de la estereocilia.
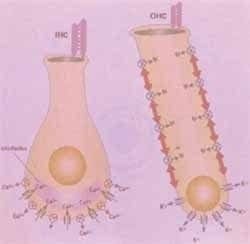 Figura 3. Transporte a través de las células ciliadas internas y externas
Figura 3. Transporte a través de las células ciliadas internas y externas
Los movimientos de la membrana timpánica, en respuesta a las ondas sonoras, son transmitidos y amplificados por la cadena oscicular y retransmitidos como ondas de compresión hacia la escala vestibular de la cóclea. Estas ondas mueven la membrana basilar causando la deflexión de las estereocilias contra la membrana tectoria. La deflexión de las estereocilias conducen a la apertura de canales iónicos que permiten la entrada de potasio al interior de la célula ciliada induciendo su despolarización. La despolarización celular genera la activación de canales de calcio conllevando a la movilización de vesículas sinápticas y posterior liberación del neurotransmisor en el espacio sináptico; de esta forma se inicia la activación del nervio coclear.
Las moléculas de miosina no convencional juegan un papel importante en el proceso de transducción manteniendo la tensión entre las uniones de los ápices de las estereocilias.
Para mantener el funcionamiento de la célula ciliada, los iones de potasio que entran en su interior debe salir, además; debe mantenerse una alta concentración a nivel de la endolinfa. Con el fin de mantener este proceso se ha descrito un mecanismo de reciclaje del potasio mediante el cual estos iones salen de la célula ciliada a nivel de su membrana basolateral por un canal de potasio, alcanzando las células de soporte del órgano de Corti. Posteriormente difunden en forma pasiva de célula a célula a través de uniones brecha (gap junctions) compuestas por una proteína multimérica, denominada conexina 26, presente en las células de soporte del órgano de Corti, células del limbo y del ligamento espiral.
Una vez los iones de potasio alcanzan la estría vascular son activamente bombeados hacia la endolinfa por canales de potasio dependientes de voltaje (2, 7, 8).
La membrana tectoria es una estructura acelular con una función mecánica en el proceso de transducción de la señal, conformada por una matriz proteica. Varios tipos de colágeno forman más de la mitad de su estructura, predominando el colágeno tipo II, y menores cantidades de tipo IX y XI. La proteína no colágeno más abundante es la a tectorina.