HLA–Cw, HLA-B, HLADRB1, HLA-DQ
Al analizar la distribución de los haplotipos conservados o extendidos, encontramos que los haplotipos conocidos en Caucásicos tuvieron una alta frecuencia en CU (0.10) en contraste con una muy baja frecuencia (0.025) en CA (p=<0.001). Solo 14 de los haplotipos del total de 450 analizados se extendieron hasta el locus A; 5 del grupo 1 hasta el A*0101, 3 del grupo 3 hasta el *2601, dos del grupo 4 hasta el *2902, 4 del grupo 5 hasta el *3002 de los Caucásicos. También, el haplotipo Africano se extiende hacia el *3001 (no mostrados en la Tabla 4).
Tabla 3. Frecuencia de bloques de DRB1 y DQB1

Tabla 4. Frecuencia de haplotipos extendidos y conservados de CMH en las poblaciones Mexicanas
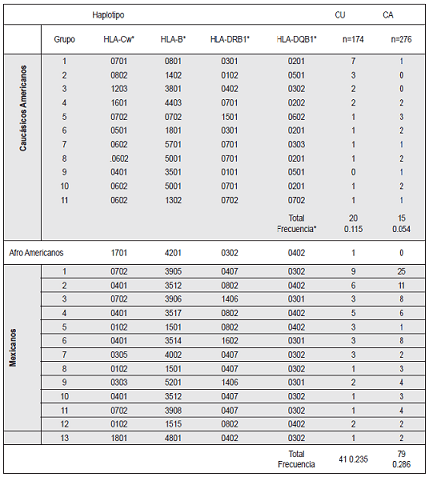
Los grupos de Caucásicos Americanos y de Africanos americanos se basan en haplotipos conocidos de esas etnias
Los haplotipos en mexicanos son hipotéticos.
* p < 0.001
Los haplotipos Mexicanos que se muestran en la Tabla 4 son hipotéticos porque para confirmarse se requiere del calculo de valores de desequilibrio de unión (LD). La frecuencia de estos haplotipos fue similar en los dos grupos (0.25 para CU vs 0.30 para CA). Los 5 primeros de los haplotipos Mexicanos en la lista tienen una frecuencia mayor del 2%. Es de resaltar que cuatro de los haplotipos en la lista, identificados con los números del 1 al 4 se extienden hasta el locus A. El primero hasta *6803 en 21 casos, el 2 hasta el *0201 en 12 casos, el 3 hasta el *2402 en 9 casos y el 4 hasta el *0206 en cinco de ellos (se contaron en ambos CU y CA).
Es importante anotar que la frecuencia de haplotipos extendidos, resultado de mezclas de Mexicanos con Caucásicos, Africanos o Asiáticos es de 15% en la población adulta y del 9% en los AC (datos no publicados). Esto indica que en el mestizaje, la frecuencia de haplotipos extendidos mestizos (combinación de dos bloques) es menor que los haplotipos Mexicanos.
Discusión
Estudios previos, basados en observaciones epidemiológicas, han propuesto que las infecciones en la infancia pueden tener un efecto posterior en la vida de los adultos (Tlaskalová-Hogenová, 2004). También es claro que las acciones de salud pública y las intervenciones médicas disminuyen las infecciones y por ende la mortalidad en poblaciones con acceso a estos servicios (Gurven, 2008).
Sin embargo, no ha sido posible determinar claramente la influencia de factores genéticos en relación con la mortalidad infantil secundaria e infecciones (Gluckman, 2004; Finch, 2004). Un reporte que soporta nuestra hipótesis mostró que la variante genética HLA-B7 es marcador de susceptibilidad para la fiebre tifoidea y la fiebre amarilla en la población de origen holandesa de Surinam (De Vries, 1979 ). Esto puede explicar porque los bloques HLA-Cw*0702, B*0702 y HLADRB1* 1501, DQB1*0602 disminuyeron en frecuencia en Surinam después de estas epidemias. De esta forma, las infecciones podrían influir en la selección natural de marcadores genéticos en la población.
Por ejemplo, aunque en este trabajo no se estudió una población infantil, creemos que los cordones umbilicales de un banco público de México pueden representar una población infantil de bajos recursos. Usando los bloques de ADN del CMH para definir el grado de mestizaje de los Mexicanos (Yunis, 2003, Yunis 2005a) encontramos diferencias en las frecuencias de los diferentes bloques entre las dos poblaciones. En particular, con respecto al DRB1*, DQB1* la frecuencia de los bloques Caucásicos en cordones umbilicales (CU) fue significativamente más alta que en los adultos (CA). Es importante anotar que el exceso de representación de este bloque en los CU se debió a que los bloques (DRB1*0301, DQB1*0201; DRB1*01501, DQB1*0602; DRB1*0402, DQB1*0302; DRB1*0401, DQB1*0302; DRB1*0701, DQB1*0201; DRB1*0401, DQB1*0301 y DRB1*0302, DQB1*0402) se encuentran aumentados en enfermedades autoinmunes como diabetes mellitus tipo-I, artritis reumatoide, lupus eritematoso sistémico, esclerosis múltiple, hipertiroidismo y enfermedad celiaca (Yunis, 2005a; Yunis, 2005b; 16; Invernizzi, 2009).
Además, la artritis reumatoide y el pénfigo vulgar en mexicanos se asocian con HLA-DRB1*0404, el cual está representado igualmente en adultos (0.08) que en CU (0.069) (Yunis, 2005a, Invernizzi, 2009). En contraste, la frecuencia de bloques Mexicanos fue mayor en las muestras de adultos que de cordones umbilicales. También consideramos que en la población mestiza mexicana, la frecuencia de haplotipos extendidos mexicanos fue mayor que la frecuencia de haplotipos mestizos (combinación de un bloque de mexicanos con un bloque de una etnia diferente, principalmente de Caucasicos). Estos hallazgos sugieren que las infecciones no tratadas adecuadamente en poblaciones de bajos recursos, con limitado acceso a sistemas de salud, pueden modular la respuesta inmune hacia un estado proinflamatorio. La modulación de la respuesta inmune asociada a alteraciones de la microbiota normal podrían causar tormentas de citoquinas, inflamación y contribuir en la mortalidad (Trifonov, 2009; WHO, 2010). De esta manera, nuestra hipótesis para el estado proinflamatorio requiere de una interacción entre la respuesta inmune, los genes del CMH y la microbiota.
Hipótesis: Mecanismo inmunitario en infecciones por genes del CMH y la microbiota en la pro-inflamación.
El periodo neonatal es crítico para la colonización del intestino y puede ser afectado por muchos factores incluyendo la edad de gestación, el ambiente al nacer, la nutrición y el uso de antibióticos (Hughes, 2009; Chow, 2009). La microbiota intestinal es la comunidad de microorganismos que participan del metabolismo intestinal y desarrollo de la inmunidad, diferenciación de células T e inflamación (Tlaskalova-Hogenova, 2005; Chow, 2009; Slack, 2009). La microbiota es regulada durante la infancia y cambios en esa población bacteriana durante el desarrollo del individuo puede modificar la intensidad de infecciones, la inflamación y, en la edad adulta, puede facilitar el desarrollo de las enfermedades autoinmunes (Invernizzi, 2009). Como esas enfermedades poligénicas tienen un componente del CMH, debemos considerar el papel de las mezclas genéticas en el desarrollo de las mismas (Awdeh, 2006; Granados, 1996; Molokhi, 2000).
La participación de infecciones en el desarrollo de autoinmunidad ha sido demostrada en el modelo murino F1(NZBxNZW) que desarrolla lupus. Al ser mantenidos en ambiente con menos gérmenes, se reduce la incidencia de lupus (Howie, 1968). Otros estudios han mostrado que las infecciones con helmintos, lactobacilo, bifidobacterias y bacterias saprofititas pueden inhibir la autoinmunidad (Mulder, 2009).
En la respuesta inmune contra agentes infecciosos participan tanto mecanismos innatos como adquiridos (Chow, 2009; Fuji, 2004; Rub, 2009). Por ejemplo, las células como macrófagos, después de fagocitar los patógenos, presentan los péptidos mediante las moléculas del CMH a las células T CD4+ (Th1) que producen IL-12 y diversas otras citocinas como IFN-, también producida por células asesinas naturales (NK). La respuesta proinflamatoria Th1 se observa primordialmente en infecciones virales y por otros patógenos intracelulares. Las células T CD4+ Th2 o son productoras de IL-4, IL-5 e IL- 13 y son importantes en el control de infecciones extracelulares (bacterias y parásitos), mientras que las células Th17 producen IL-17, IL-21 y/o IL-22 y controlan infecciones extracelulares (hongos y bacterias). Por otro lado, las células T reguladoras (Tregs), productoras de IL-10 y TGF-, participan de la respuesta a esos patógenos mediante su acción anti-inflamatoria (Okada, 2010; Zelate, 2009; Awasthi, 2009; Ivanov, 2008; Calcinaro 2005; Torchinsky, 2009; Kastelein, 2007).
La microbiota también interactua con células Th17 del intestino y con las Tregs a través de factores de trascripción RoRt y FoxP3, respectivamente (Chow, 2009). Sin embargo, no se conoce la participación de la microbiota en el estado proinflamatorio acentuado, ni el papel de la IL-27 en relación al grado de proinflamación, aunque se sabe que IL-27 reduce el número de células T regs (Kastelein, 2007). Consideramos que las investigaciones futuras que evalúen el grado de inflamación, demostrarán cambios en la capacidad funcional de las células Tregs y en las células Th17 (Chow, 2009) así como la relevancia de la sobreproducción de IL-12 e IL-27 en individuos con alteraciones del equilibrio de microbiota, especialmente a aquellos que poseen genes del CMH (clase II) de susceptibilidad a enfermedades autoinmunes.
Agradecimientos
El estudio fue apoyado por el Departamento de Cancer immunology and AIDS del Dana Farber Cancer Institute y por una donación del Dr. Julio Granados.
El contenido de este artículo fue presentado en gran parte durante el Congreso Médico “Bicentenario de la Independencia de Colombia”, Academia Nacional de Medicina y Asociación de Médicos Colombianos en Estados Unidos-USMA 30 años.
Referencias
1. Awasthi A, Kuchroo VK. Th17 cells: from precursors to players in inflammation and infection. International Immunology. 2009; 21; 489-98.
2. Awdeh ZL, Yunis EJ, Audeh MJ, Fici D, Pugliese A, Larsen CE et al., A genetic explanation for the rising incidence of type 1 diabetes, a polygenic disease. Journal of Autoimmunity. 2006; 27:174-180.
3. Calcinaro F, Dionisi S, Marinaro M, Candeloro P, Bonato V, Marzottiet S, et al., Oral probiotic administration induces IL-10 production and prevents spontaneous autoinmune diabetes in the NOD Mouse. Diabetologia 2005; 48: 1565-1575.
4. Cantú de León D, Pérez-Montiel D, Villavicencio V, García Carranca A, Mohar Betancourt A, Acuña-Alonzo V, et al., High resolution human leukocyte antigen (HLA) class I and class II allele typing in Mexican mestizo women with sporadic breast cancer: case-control study. BMC Cancer. 2009; 9:48.
5. Collins D and Collins J. Maternal and infant health in diverse settings. American Journal of Public Health. 2007; 97; 1191-1197.
6. Chervonsky AV. Influence of microbial environment on autoimmunity. Nature Immunology 2010; 11: 28-35.
7. Chow J and Sarkis MK. Getting the Bugs out of the Immune System: Do Bacterial Microbiota “Fix” Intestinal T Cell Responses? Cell Host & Microbe. 2009; 51: 8-12.
8. Hughes SM, Amadi B, Mwiya M, Nkamba H, Tomkins A, Goldblatt D. Dendritic cell anergy results from endotoxemia in severe malnutrition, Journal of Immunology. 2009; 183: 2818-26.
9. De Vries RR, Meera P Khan P, Bernini LFE, van Loghem F, Van Rood. JJ. Genetic control of survival in epidemics. International Journal of Immunogenetics. 1979; 6: 271-287.
10. Finch CE, Crimmins EM. Inflammatory exposure and historical changes in human life-Spans. Science. 2004; 305: 1736-1739.
11. Fujii S, Liu K, Smith C, Bonito AJ, Steinman RM. The linkage of innate to adaptive immunity via maturing dendritic cells in vivo requires CD40 ligation in addition to antigen presentation and CD80/86 costimulation. Journal of Experimental Medicine. 2004; 199: 1607-18.
12. Gluckman PD, Hanson MA. Living with the past: evolution, development, and patterns of disease. Science. 2004; 305: 1733-173.
13. Granados J, Vargas-Alarcon G, Andrade F, Melin-Aldana H, Alcocer-Varela J, Alarcon de Segovia D. The role of HLA-DR alleles and complotypes through the ethnic barrier of systemic lupus erythematosus in Mexicans. Lupus. 1996; 5:184-189.
14. Gurven M, Kaplan HW, Finch CE, Crimmins EM. Aging and Inflammation in Two Epidemiological Worlds, Journal of Gerontology: Medical Sciences. 2008; 63A. 000-000.
15. Howie JB, Helyer BJ. The immunology and pathology of NZB mice. Advances in Immunology. 1968; 9:215-266.
16. Invernizzi P, Gershwin ME. The genetics of human autoimmune disease. Journal of Autoimmunity. 2009; 33: 290-299.
17. Ivanov II, Frutos R, Manel N, Yoshinaga K, Rifkin DB, Balfour Sartor, et al., Specific microbiota direct the differentiation of IL-17 producing T-helper cells in the mucosa of the small intestine. Cell Host Microbe. 2008; 4: 337-349.
18. Kastelein RA, Hunter CA, Cua DJ. Discovery and biology of IL- 23 and IL-27: Related but functionally distinct regulators of inflammation. Annual Reviews of Immunology. 2007; 25: 221–42.
19. Miller SA, Dykes DD, Polensky HF. A simple salting out procedure for extracting DNA from human nucleated cells. Nucleic Acids Research. 1998; 11: 12-15.
20. Molokhi M, McKeigue P. Risk for rheumatic disease in relation to ethnicity and admixture. Arthritis Research. 2000; 2:115-125.
21. Mulder IE, Schmidt B, Stokes CR, Lewis M, Bailey M, Aminov RI, et al., Environmentally-acquired bacteria influence microbial diversity and natural innate immune responses at gut surfaces. BMC Biology. 2009; 7:79.
22. Okada H, Khun C, Fellet H, Bach JF. The hygiene hypothesis and allergic diseases: an updte. Clinical and Experimental Immunology. 2010; 160: 1-9.
23. Rub A, Dey R, Jadhav M, Kamat R, Chakkaramakkil S, Majumdar S et al., Cholesterol depletion associated with Leishmania major infection alters macrophage CD40 signalosome composition and effector function. Nature Immunology. 1009; 10: 273-280.
24. Semenza JC. Strategies to intervene on social determinants of infectious diseases. Euro Surveill. 2010; 15: 19611 https://www. eurosurveillance.org/ViewArticle.
25. Slack E, Hapfelmeier S, Stecher S, Velykoredko Y, Stoel M, Lawson ME, et al., Innate and adaptive immunity cooperate flexibly to maintain host-microbiota mutualismo. Science. 2009; 325: 617-620.
26. Tlaskalová-Hogenová H, Stepánková R, Hudcovic T, Tucková L, Cukrowska B, Lodinová-Zádníková R, et al., Commensal bacteria (normal microflora), mucosal immunity and chronic inflammatory and autoimmune diseases. Immunology Letters 2004; 93:97–108.
27. Tlaskalova-Hogenova H, Tuckova L, Mesteckyy J, Kolinskaz J, Rossmann P, Stepankova R et al., Interaction of mucosal microbiota with the innate immune system. Scandinavian Journal of Immunology. 2005; 62 (Suppl. 1): 106–113.
28. Torchinsky MB, Johan G, Andrea MP, Magarian BJ. Innate immunity recogniton of infected apoptotic cells directs TH17 cell differentiation. Nature. 2009; 458: 78-82.
29. Trifonov V, Khiabanian H, Rabadan R. Geographic Dependence, Surveillance, and Origins of the 2009 Influenza A (H1N1) Virus. New England Journal of Medicine. 2009; 61:115–119.
30. Writing Committee of the WHO Consultation on Clinical Aspects of Pandemic (H1N1) 2009 Influenza Clinical Aspects of Pandemic 2009 Influenza A (H1N1) Virus Infection. World Health Organization New England Journal of Medicine. 2010; 1708–19.
31. Yunis EJ, Larsen CE, Fernandez-Viña M, Awdeh ZL, Romero T, Hansen JA et al., Inheritable variable sizes of DNA stretches in the human MHC: conserved extended haplotypes and their fragments or blocks. Tissue Antigens. 2003; 62: 1-20.
32. Yunis EJ, Larsen CE, Fernandez-Viña M, Awdeh ZL, Cao K, Romero T, Alper CA, Hansen JA. Human MHC haplotypes and their fragments or blocks. In: Proceedings of the 13th International Histocompatibility Workshop. Oxford: Blackwell Munksgaard. 2005a.
33. Yunis EJ, Zuniga J, Larsen CE, Fernandez-Vina M, Granados J, Awdeh ZL, Alper CA. Single nucleotide polymorphism blocks and haplotypes: human MHC block diversity. In Encycopedia of Molecular Cell Biology and Molecular Medicine. Volume 13. Meyers RA. Weinheim: Wiley-VCH. 191-215. 2005b.
34. Zelate T, De Luca A, D’Angelo C, Moretti S, Romani L. Th17 in host defense. European Journal of Immunology. 2009; 39: 634-675.