Capítulo 5
Anatomía e Histología de los Islotes Pancreáticos

El páncreas (palabra griega que significa todo carne) es una glándula elongada anexa al aparato digestivo, anidada alrededor del duodeno, con una función digestiva exocrina y una función endocrina basada en los islotes de Langerhans (Figura 1).
De los cincuenta a setenta y cinco gramos que pesa el páncreas, sólo un gramo corresponde a tejido insular; en la histología convencional, los islotes se ven como grupos de células relativamente pálidas diseminadas por el tejido exocrino que se tiñe de oscuro.
El número de islotes va de un cuarto de millón a un millón y tres cuartos, su diámetro aproximado es de ciento cincuenta micras y son más numerosos hacia la cola del páncreas, aunque se encuentran distribuidos por todo el órgano. Hay tres tipos de células:
- El sesenta a noventa por ciento de las células corresponden al tipo beta, productoras de insulina
- La mayoría de las demás son alfa, productoras de glucagón
- Otro pequeño porcentaje está representado por las delta, productoras de somatostatina.
Las células beta ocupan la parte central del islote y están rodeadas por las alfa y delta. Los islotes –que producen otras hormonas en cantidades muy pequeñas- son altamente vascularizados pues reciben proporcionalmente cerca de diez veces más flujo sanguíneo que el páncreas exocrino.
El sistema nervioso autónomo inerva las células insulares y modula la secreción hormona a través de señales simpáticas o para-simpáticas.
Insulina
Química y Regulación

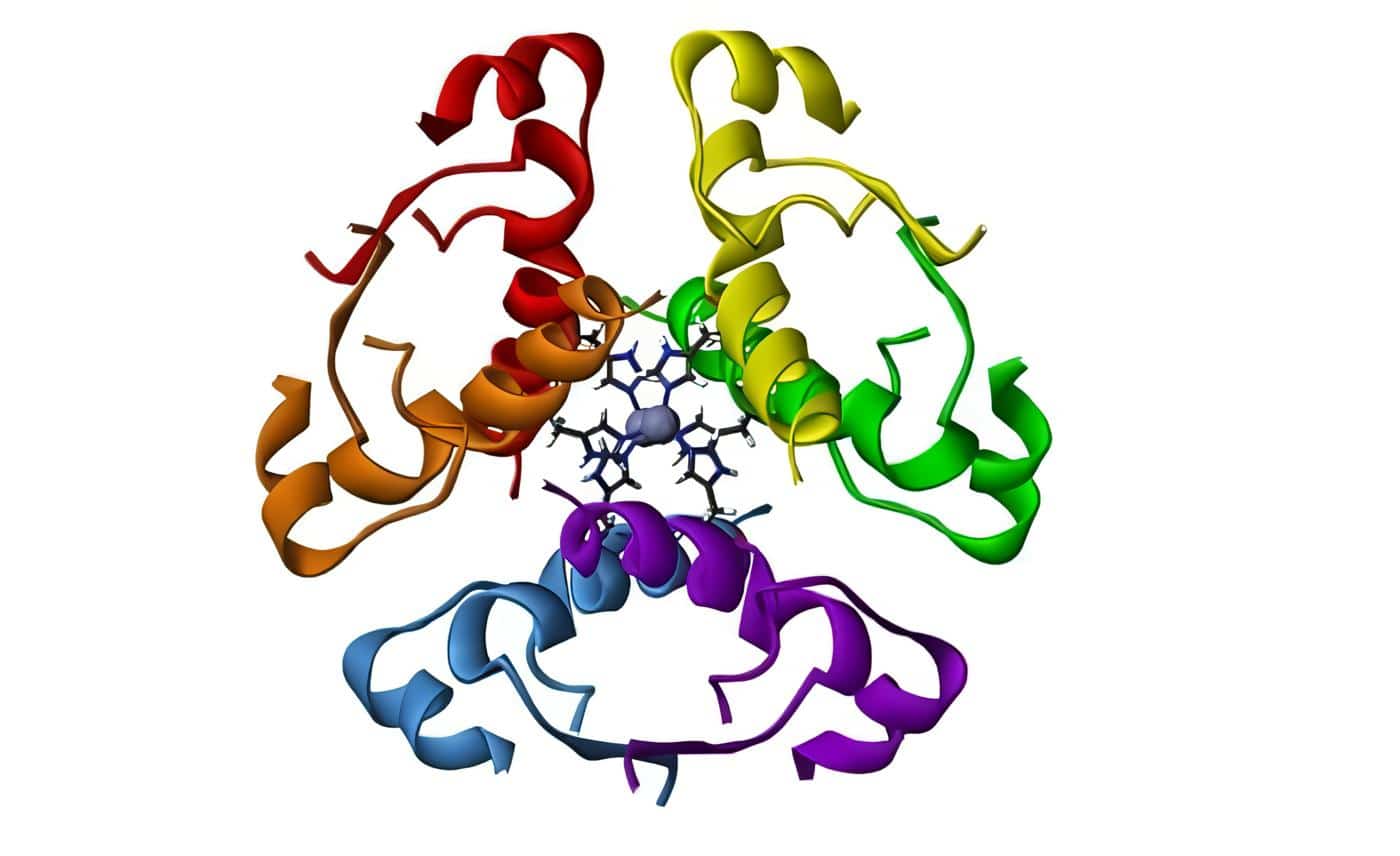
La insulina es un polipéptido que contiene cincuenta y un aminoácidos, está compuesto por las cadenas A y B unidas entre sí por dos puentes disulfídicos; la cadena A tiene veintiún aminoácidos y la B, treinta, y se encuentran dispuestas en forma de hélice, lo cual influye en las propiedades físico-químicas de la hormona (Figura 2).
En la cadena A hay también un anillo disulfídico que rodea seis aminoácidos en la misma forma que ocurre en los polipéptidos vasopresina y ocitocina por lo que se cree que este anillo sea indispensable para la fijación al receptor; los puentes disulfídicos también son indispensables para la actividad biológica de la insulina.
La secuencia de aminoácidos se conserva bastante entre los diferentes vertebrados por lo que la insulina producida por el páncreas de alguno de estos vertebrados puede utilizarse en otro, como se utilizó en humanos en las primeras décadas después del descubrimiento de la hormona.
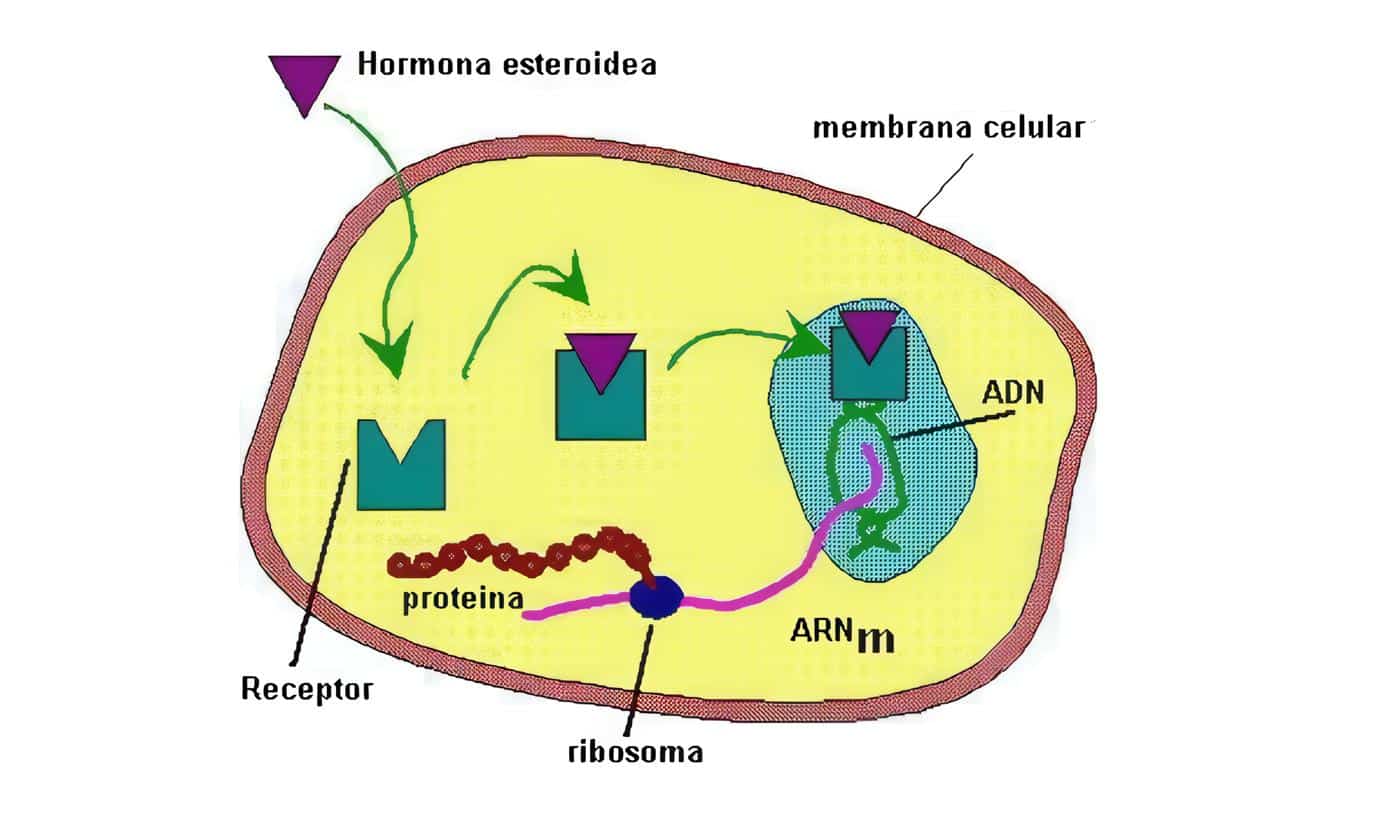
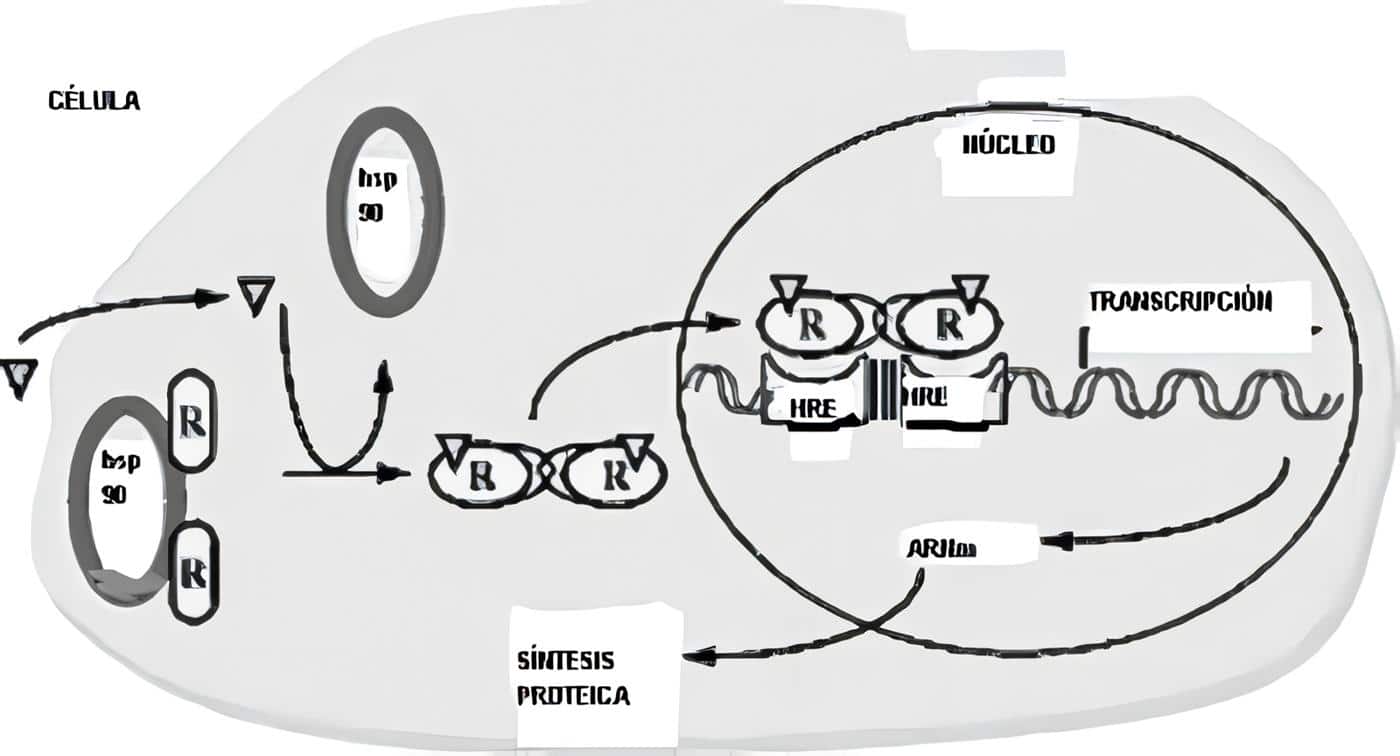
El precursor proteico de la hormona – de mayor peso molecular- se llama Pro insulina; el ARN mensajero de la insulina se traduce como un precursor de una sola cadena –la pre-proinsulina- que al insertarse en el retículo endoplásmico da lugar a la pro insulina.
Esta última –bajo la acción de peptidasas- se desdobla, dando lugar a insulina y al péptido de conexión (péptido C), sin acción biológica, pero que sirve de marcador en la secreción de insulina; ambos son depositados en gránulos, en el aparato de Golgi. Cuando la célula beta recibe un estímulo apropiado, la insulina se libera por exocitosis y pasa a la sangre, al igual que el péptico C (Figura 3).
(Lea También: Transporte, Receptor y Degradación de los Islotes Pancreáticos)
 Se calcula que el páncreas produce diariamente unas cincuenta unidades de insulina, cuyo principal estímulo secretor es la glucosa.
Se calcula que el páncreas produce diariamente unas cincuenta unidades de insulina, cuyo principal estímulo secretor es la glucosa.
 Se calcula que el páncreas produce diariamente unas cincuenta unidades de insulina, cuyo principal estímulo secretor es la glucosa.
Se calcula que el páncreas produce diariamente unas cincuenta unidades de insulina, cuyo principal estímulo secretor es la glucosa.Cuando este carbohidrato se administra endovenosamente, los niveles insulínicos suben rápidamente en sangre pero no llegan a ser tan altos como cuando la glucosa se administra por vía yeyunal a través de un catéter; esto es debido a las hormonas intestinales o incretinas, el péptido insulinotrópico dependiente de glucosa, también denominado péptido inhibitorio gástrico o “enterogastrona” originalmente (GIP) y el péptido glucagón-símil 1 (GLP-1), secretagogos que estimulan la producción de insulina.
Existen estímulos secundarios: cuando este carbohidrato se administra endovenosamente, los niveles insulínicos suben rápidamente en sangre pero no llegan a ser tan altos como cuando la glucosa se administra por vía yeyunal a través de un catéter.
Esto se debe a la secreción –cuando se come- de dos hormonas intestinales insulinotrópicas llamadas incretinas, el péptido tipo glucagón (GLP-1) y el péptido inhibitorio gástrico (GIP), también denominado péptido insulinotrópico dependiente de glucosa (inicialmente llamado enterogastrona); estos secretagogos estimulan la producción de insulina.
La exenatida y la liraglutida son nuevos análogos incretino-miméticos. Además de su acción insulinotrópica, el GLP-1 inhibe la secreción de glucagón, baja los niveles de glicemia, promueve la diferenciación de las células progenitoras de los islotes y mejora la función y la duración de la beta-célula.
Durante el ayuno, la insulinemia esta por debajo de lo normal pero después de la ingestión de alimentos sus niveles se elevan por encima de lo normal.
La principal sustancia que estimula la secreción de insulina es la glucosa. Las etapas involucradas en este proceso son las siguientes.
- Un transportador de glucosa –por difusión facilitadora- introduce el monosacárido a la beta célula.
- Al aumentar la concentración de glucosa dentro de la célula beta, la membrana se despolariza e ingresa calcio proveniente del espacio extra-celular, lo que induce la exocitosis de los gránulos de insulina
- El aumento en la glucosa intra-celular activa también mecanismos calcio-independientes de la secreción de insulina.
La secreción insulínica es bifásica. Cuando se logra experimentalmente durante una hora un incremento de dos a tres veces los niveles basales de ayuno en una persona (80 a 90 mg/dl), los niveles de insulina aumentan marcadamente, debido a la liberación de insulina ya formada y almacenada.
Después comienza un segundo pico de insulinemia, que corresponde a nueva hormona sintetizada que se va liberando de inmediato, lo que indica que la hiperglicemia post-prandial no sólo estimula la liberación de insulina almacenada sino también la trascripción nuclear del gen insulínico y la traducción de su ARN mensajero.
Otros factores que regulan la secreción insulínica –aunque algunos de ellos son poco activos- se pueden ver en la tabla 1.
TABLA 1
Factores que regulan la secreción de insulina
ESTIMULAN |
INHIBEN |
| Glucosa | Diuréticos tiazídicos |
| Fructosa, otros monosacáridos | Diazóxido |
| Aminoácidos (arginina, leucina) | Beta-bloqueadores |
| Ácidos grasos | Difenil-hidantoinato sódico |
| Estimulantes adrenérgicos beta 2 :Salbutamol | Puromicina |
| Sulfonilureas | Estimulantes adrenérgicos alfa: Norepinefrina |
| Agonistas colinérgicos | Somatostatina |
| Estimulación vagal | Colchicina |
| Xantinas (teofilina) | Insulina |
| Gastrina | Vagotomía |
| Secretina | 2- desoxi-glucosa |
| Glucagón | Manoheptulosa |
| Prostaglandinas | l-asparaginasa |
| Hormonas contra-reguladoras de la insulina (indirectamente) |
Ayuno |
| Biguanidas |