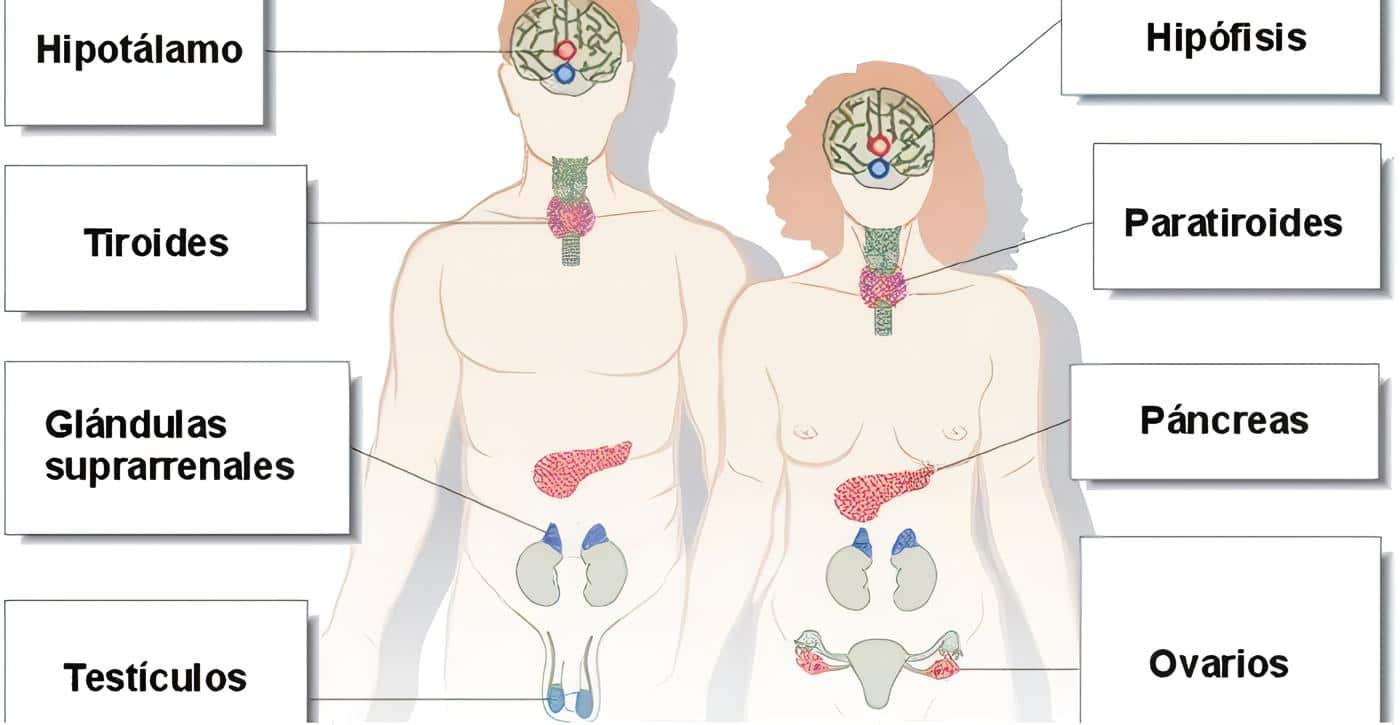
Funciones hormonales
Cada hormona puede tener funciones diferentes, aunque a menudo algunas son tal vez más importantes que otras. La acción puede ser directa –en los tejidos- o a través de la liberación de otras, como en el caso de las hormonas tróficas. En la tabla 2 podemos ver las más importantes.
Tabla 2 (De Raff, modificado)
Funciones generales que regulan las hormonas
Depósito, distribución y gasto de calorías, termogénesis
>Reproducción |
>Ciclo menstrual, ovulación, espermatogènesis, embarazo y lactancia |
|---|---|
| >Crecimiento y Desarrollo | Diferenciación sexual, características sexuales secundarias, aumento Masas ósea y muscular, velocidad del crecimiento linear |
| Homeostasis o Equilibrio del Medio Interno | Volumen extracelular, tensión arterial, equilibrio electrolítico y de Iones plasmáticos como el calcio, mantenimiento de las reservas Energéticas, como grasas, glicógeno, proteínas. |
| Suministro de Energía | Depósito, distribución y gasto de calorías, termogénesis |
| Conducta | Ingestión de agua y comida, libido y estado de ánimo. |
Control de la secreción hormonal
Las hormonas hidrosolubles se pueden clasificar (según Koeslag) en tres diferentes categorías.
A) Las hormonas que ejercen contrarregulaciòn de otras –control con riendas- o reguladoras del medio interno.
B) Reguladoras balísticas, en las que los mensajes que envían no son luego confirmados (son de asa abierta) pues se generan en situaciones especiales o de emergencia.
C) Mensajeros de relevo de hormonas, generalmente liposolubles, (que son de asa cerrada).
Insulina y el glucagón
Entre las primeras, o que ejercen entre sí una contrarregulación están la insulina y el glucagón (que regulan la glicemia), la PTH y la calcitonina (que regulan el calcio ionizado), la ADH y el ANP (que regulan osmolalidad, volumen sanguíneo, control del sodio), la EPO que controla la oxigenación de la hemoglobina, pero que no tiene una hormona contrarreguladora, y la gastrina y la secretina, que responden a la acidez gastroduodenal.
Reguladoras balísticas son la epinefrina, que de urgencia aumenta la glicemia y la tensión arterial, la ocitocina, que produce eyección làctea y contracción uterina, la prolactina, que regula producción de leche y la fertilidad, la CCK y las interleucinas.
En cuanto a los mensajeros de relevo están las hormonas hipotalàmicas, las adenohipofisiarias u hormonas tròficas en general, la angiotensina II que regula la secreción de aldosterona y la PTH, que regula la l-alfa hidroxilaciòn renal de la vitamina D3, generando el calcitriol.
Las hormonas circulantes interactúan con las células blanco en mayor o menor grado de acuerdo a su concentración plasmática y a su afinidad por el receptor principalmente. La concentración depende de la velocidad de síntesis, liberación, degradación y eliminación.
Un mecanismo de control fisiológico común es la servo-regulación (que puede semejarse a la forma como funciona un calentador central, cuyo termostato actúa a una determinada temperatura ambiente, apagándolo o encendiéndolo. La retroalimentación (“feed-back”) más común en fisiología endocrina es la negativa en los diferentes ejes (hipotálamo-hipófisis con suprarrenales, gonadas o tiroides), que a veces puede ser positiva (estradiol y LH).
Los circuitos pueden ser largos (hipófisis a glándula periférica), cortos (hipotálamo a hipófisis) o ultracortos.
La retroalimentación puede también operar por medio de la concentración plasmática de un sustrato; tal es el caso de la insulina y del glucagòn que se segregan en respuesta a un nivel dado de glicemia, o la parathormona y la calcitonina, que lo hacen de acuerdo a los niveles de calcio ionizado.
Las hormonas se segregan de manera pulsátil, con pulsos cortos (de forma que logran evitar la regulación de receptores hacia abajo), o largos, de acuerdo a ritmos circadianos.
¿Cómo actúan las hormonas?
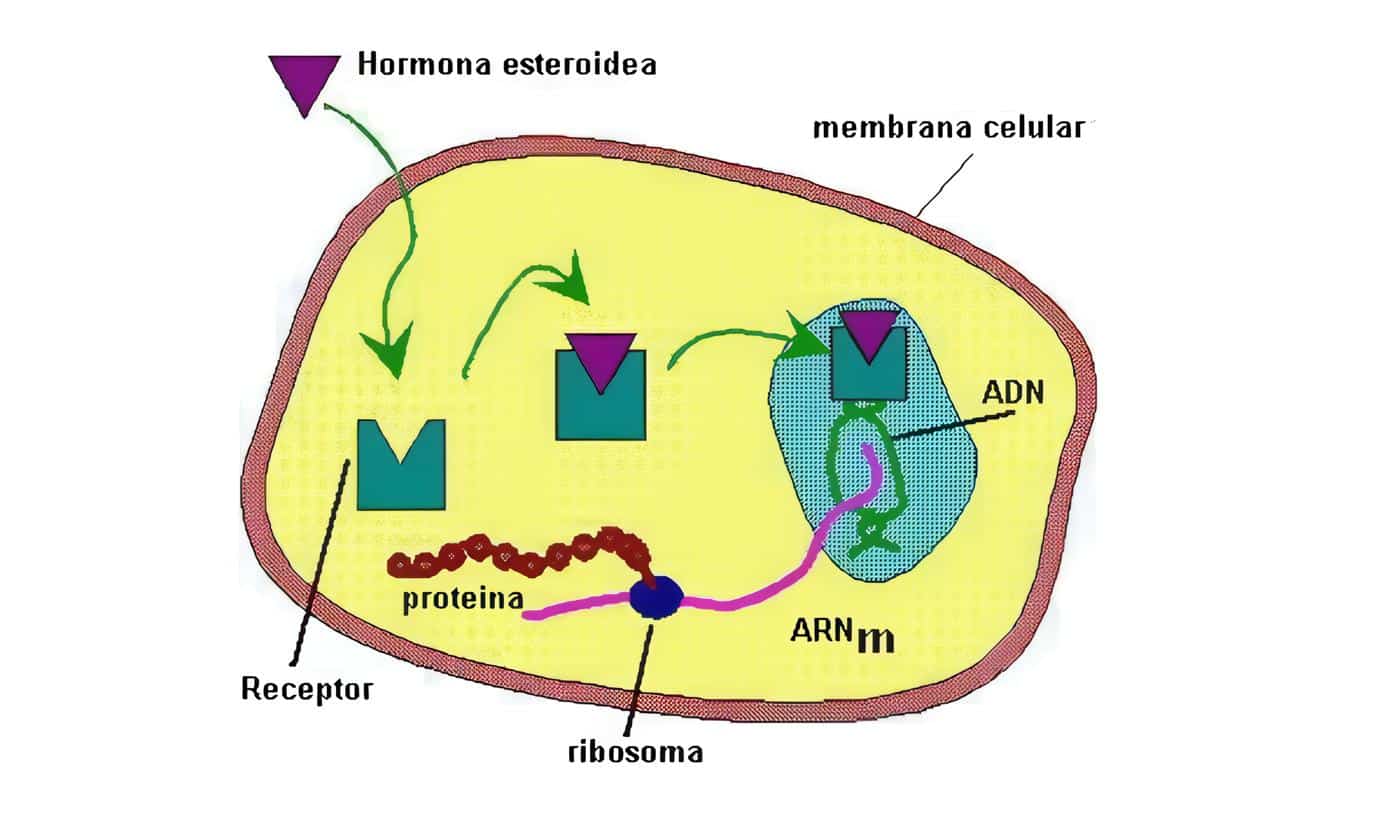
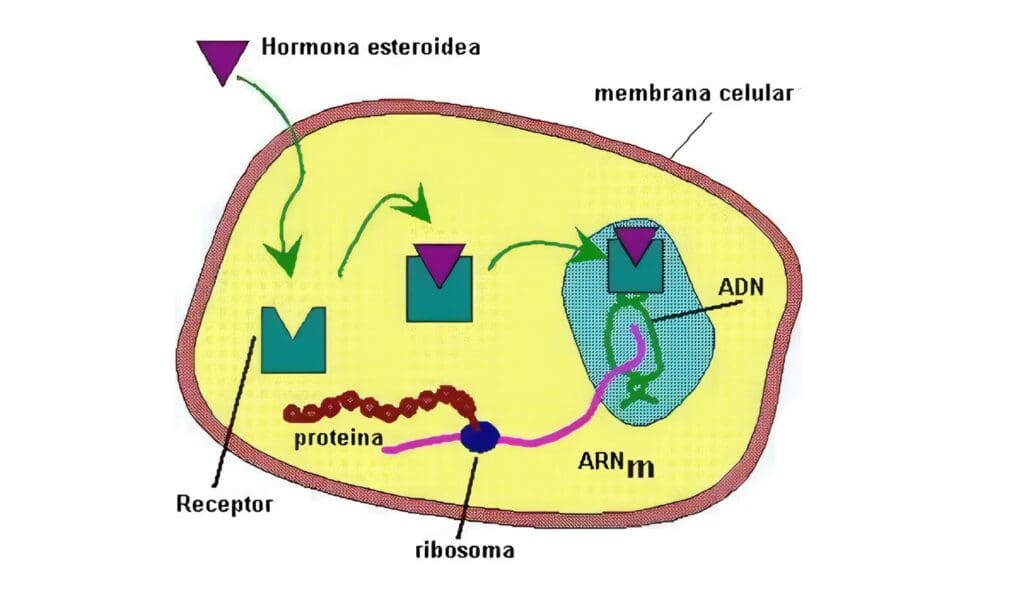
La forma como las hormonas actúan en las células blanco es por medio de activaciones enzim´sticas o por modulación de la expresión genética, estimulando la trascripción de un grupo específico de genes.
Esta actividad la logran por medio de la interacción, bien con receptores de membrana, o bien con receptores nucleares, según que la molécula sea hidrosoluble o liposoluble, lo que induce un cambio en la estructura del receptor.
La biología molecular y la ultraestructura nos muestran entonces que la morfología celular y la de sus componentes y organelas no permanecen estáticas, sino que cambia continuamente con los procesos fisiológicos.
Aunque por razones didácticas se ha tratado de esquematizar la interacción de las moléculas señaladoras y sus receptores, estas acciones son complejas, tanto porque los receptores usualmente pertenecen a “superfamilias” con centenares de miembros, como porque los procesos comprometen innumerables proteínas, enzimas, iones y actividades genéticas.
Como sabemos, las membranas celulares están compuestas de lípidos, proteínas y carbohidratos. La bicapa lipídica está integrada por fosfolìpidos con una cabeza polar hidrofílica y una cola hidrofóbica ubicada en la parte central o media de la membrana; la composición varía en los aspectos internos y externos de la membrana celular lo que lleva a una asimetría: la parte externa contiene fosfatidil colina y esfingomielina (además de colesterol), mientras que la interna tiene fosfatidil serina, etanolamina e inositol.
Estos fosfolípidos son móviles de manera lateral o transversa.
Las proteínas de membrana son integrales (o sea que atraviesan la bicapa), o periféricas (bien en la parte externa o en la interna), y pueden ser móviles o estar fijas. Los carbohidratos se encuentran en la parte externa de la membrana plasmática, constituyendo glicoproteínas y glicolípidos, formando una capa conocida como glucocáliz, que sirve para fijar el calcio extracelular y de matriz de fijación para otras células.
El paso de sustancias a través de la membrana se hace por medio de mecanismos de difusión facilitada (simple o por medio de canales), o por transporte activo, cuando la difusión es mediada por moléculas portadoras. El oxígeno, el dióxido de carbono e incluso el agua, atraviesan la capa lipídica con facilidad, pues son lipófilas y sin carga.
La glucosa, los aminoácidos y los iones, requieren de canales o de proteínas que ayudan al transporte hacia el interior de la célula. Los transportes activos pueden ser primarios (como en el caso de la bomba de sodio/potasio) o secundarios; entre los últimos está el cotransporte, por una proteína energizada (Vg. Absorción de azúcares y aminoácidos en el aparato digestivo), o contra-transporte (intercambio de sodio por hidrógeno y por calcio).
Receptores de membrana y segundos mensajeros.
Los receptores en la membrana celular tienen regiones que contribuyen a tres dominios básicos, como son los extracelulares (o dominios que se fijan a los ligandos); los transmembrana (que incluyen aminoácidos hidrofóbicos que se insertan en la bicapa lipìdica y anclan el receptor en la membrana); muchos de ellos (pero no todos) atraviesan siete veces la membrana; y finalmente están los dominios intracelulares (colas o asas intracitoplásmicas de los receptores), que son la región verdaderamente efectora hormonal, pues hay activación de proteínas cinasas, las que fosforilan proteínas, generalmente enzimas.
Estos receptores de membrana son de varias clases, siendo los más comunes los siete transmembrana acoplados a proteínas G (llamados 7TM o GPRC), proteínas que al ser activadas producen los segundos mensajeros, que actúan como señales intracelulares cuando la hormona no puede atravesar la membrana, como en el caso de las proteicas.
Estos segundos mensajeros son:
1) El adenosín monofosfato cíclico (AMPc).
2) El 1, 4,5-inositol trifosfato (IP3), el diacilglicerol (DAG) y el calcio, que podríamos decir pertenecen al sistema fosfoinosìtidos-calcio.
3) El guanosín monofosfato cíclico (GMPc). Una hormona puede utilizar diferentes segundos mensajeros y estos a su vez pueden ser producto de la interacción de los receptores con diferentes hormonas.
 Como anteriormente decíamos, los receptores de membrana usualmente- pero no siempre- son los 7TM o GPCR, acoplados a proteínas G; pertenecen a una superfamilia que cuenta alrededor de mil miembros, entre los cuales están los adrenérgicos, colinérgicos, dopaminérgicos, histaminérgicos, receptores de prostaglandinas, y receptores de péptidos del tipo vasopresina, ocitocina y angiotensina, de proteínas como glucagón, FSH, LH y TSH, además de otros GPCRs en lengua, nariz y retina que pueden fijar moléculas con sabor, olor o luminosidad.
Como anteriormente decíamos, los receptores de membrana usualmente- pero no siempre- son los 7TM o GPCR, acoplados a proteínas G; pertenecen a una superfamilia que cuenta alrededor de mil miembros, entre los cuales están los adrenérgicos, colinérgicos, dopaminérgicos, histaminérgicos, receptores de prostaglandinas, y receptores de péptidos del tipo vasopresina, ocitocina y angiotensina, de proteínas como glucagón, FSH, LH y TSH, además de otros GPCRs en lengua, nariz y retina que pueden fijar moléculas con sabor, olor o luminosidad.
Este es el caso del receptor visual de luz – la rodopsina- cuya señal de transducción nos permite ver en la penumbra (visión de bastoncitos) o el color (visión de conos); diferentes rodopsinas absorben la luz de diversas longitudes de onda en forma máxima; la proteína G que allí se activa se llama transducina.
(Lea También: Receptores Intracelulares)
Estas proteínas G, fijadoras de guanina-nucleótido, son heterotriméricas, y tienen tres sub-unidades:
Alfa (la que es activa, está ligada a guanosín-difosfato o GDP y tiene actividad de GTPasa), beta (inactiva, de las que hay cinco conocidas) y gamma (diez conocidas); las sub-unidades a su vez tienen sub-familias.
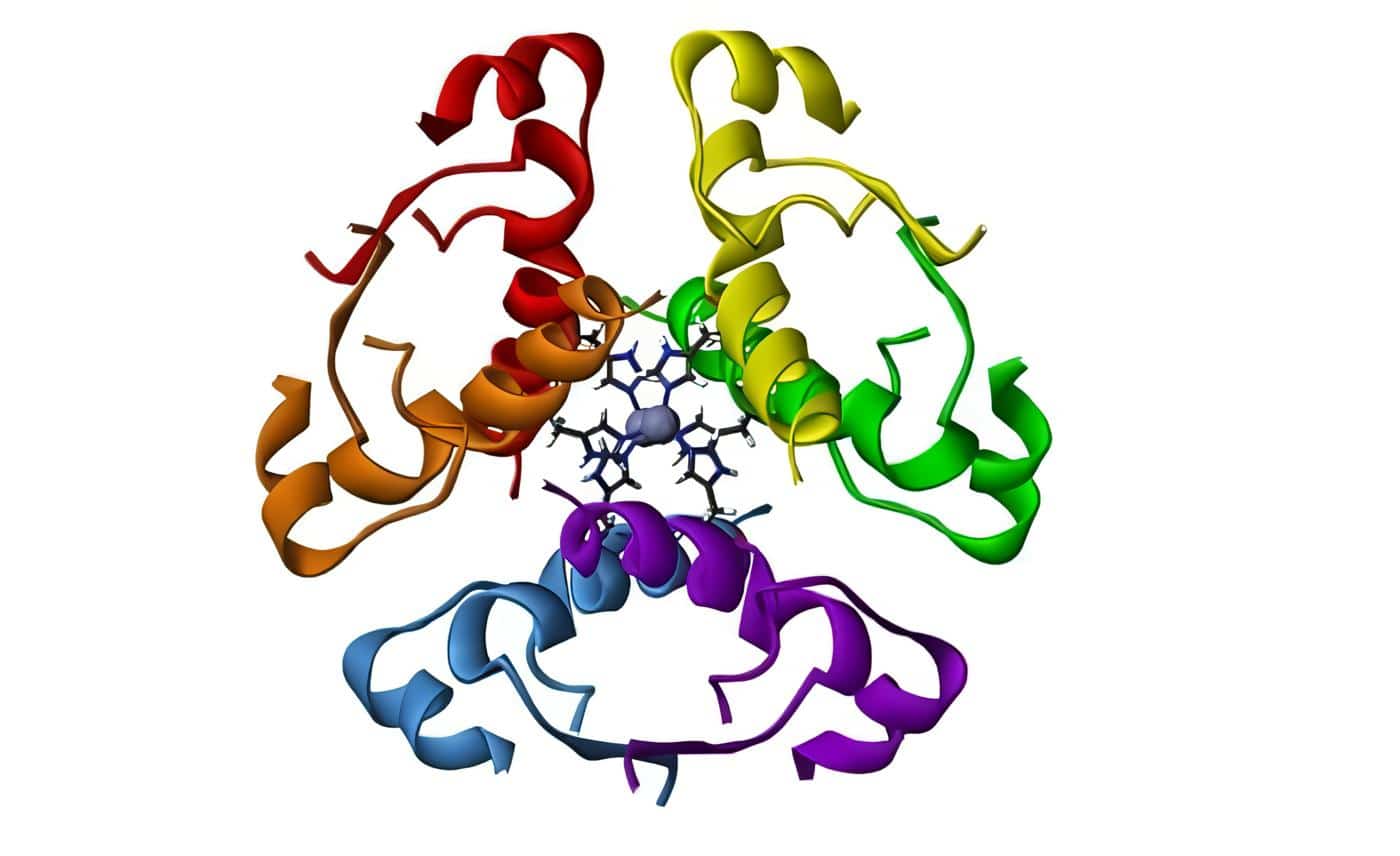
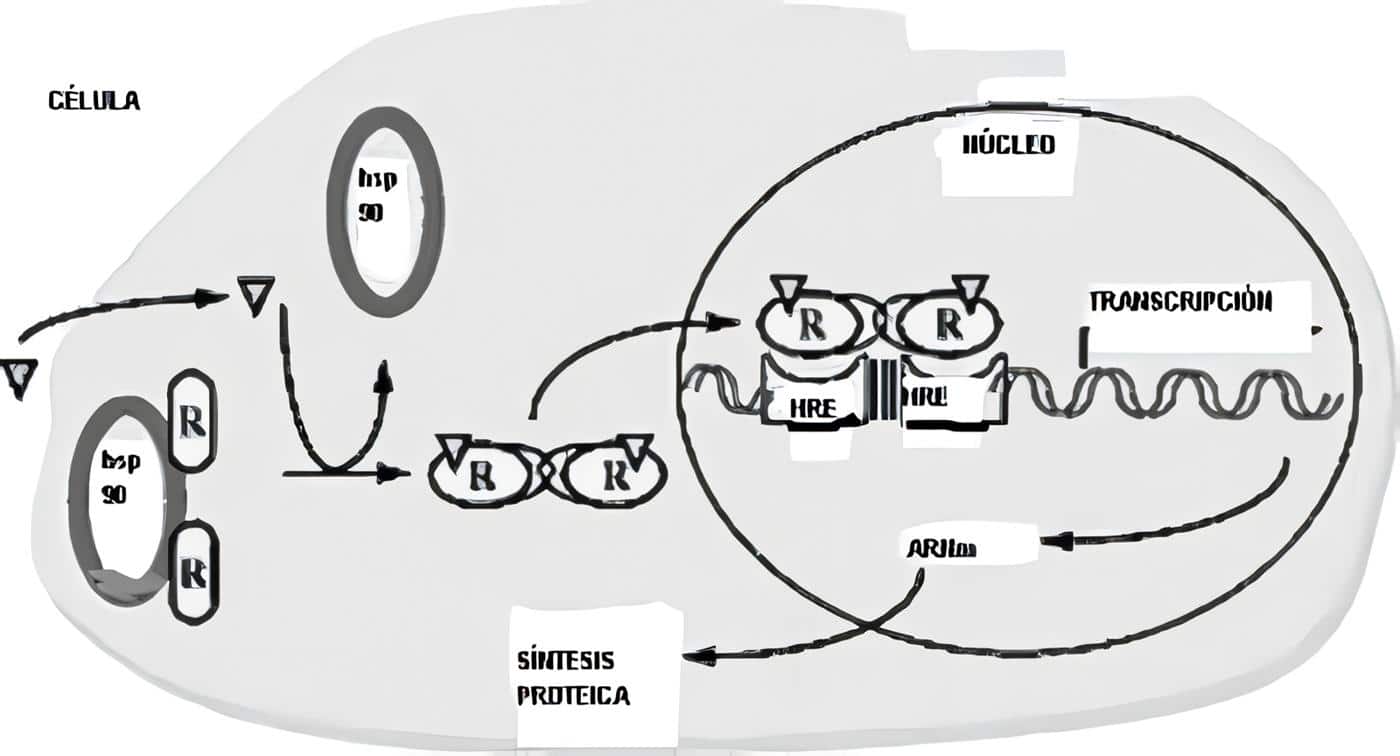
Las sub-familias de la sub-unidades alfa son: la Gs (estimulante, precisamente la que es blanco de la toxina liberada por el Vibrio cholerae), que activa la adenil-ciclasa, aumenta el AMP cíclico que a su vez activa la proteína cinasa A; enseguida se produce la activación del factor CREB de trascripción (similar al HRE nuclear para los esteroides), que es la proteína fijadora del elemento de respuesta del cAMP; este CREB se liga a su elemento de respuesta en los promotores de los genes que responden a la hormona, lo que inicia la trascripción.
La secuencia de los eventos de la activación de las Gs comienza con la fijación del ligando al receptor, lo que al producir un cambio alostèrico o de conformación de la estructura terciaria de la proteína G, permite que la GDP ligada a la sub-unidad alfa sea desplazada por GTP; este, al ligarse, disocia la sub-unidad alfa, que se fija entonces a la adenil-ciclasa y la activa, lo que genera AMPc (Figura 1).
El GTP es hidrolizado por una GTPasa, lo que permite que la sub-unidad alfa deje la adenil-ciclasa y se reasocie con las beta y gama. Las Gs están asociadas con los receptores de hormonas tales como los beta-adrenèrgicos, ACTH, glucagòn, LH, FSH, TSH, PTH y vasopresina (acción renal).
El caso clásico de la acción del segundo mensajero AMPc y la cascada subsiguiente es la activación de la fosforilasa hepática (por acción de la epinefrina y del glucagòn), que desdobla el glicógeno para producir glucosa 1 monofosfato, luego glucosa 6 fosfato que por acción de la glucosa 6 fosfatasa da lugar a glucosa, liberada a la sangre por el hígado. El AMPc se metaboliza por la acción de la fosfodiesterasa.
Otros receptores se acoplan con la proteína G-alfa-q (o Gq) que activa la fosfolipasa C beta, produciendo IP3 y DAG. El primero libera directamente calcio intracelular al citosol, cuando interactúa con su receptor, que es entonces fosforilado; el DAG, que activa la proteína cinasa C y puede actuar sinérgicamente con el IP3, reduciendo la concentración de calcio necesaria para activar la cinasa.
El DAG y el calcio modulan la actividad de la proteína cinasa a través de la calmodulina; esta pertenece a una sub-clase de proteínas que fijan calcio, que activa una proteína fosfatasa, la calcineurina, enzima muy importante en la activación de los linfocitos T, en la hipertrofia muscular y en la memoria retrógrada.
Como el mantenimiento de niveles bajos de calcio citosólicos es esencial para el buen funcionamiento celular, esto se logra entre otras cosas por el sistema calmodulina/ calcineurina que intervienen en actividades de fosforilación y defosforilaciòn de proteínas cinasas que son blanco del receptor de IP3.
La Gq se acopla a receptores como los de vasopresina (acción vascular), TRH, GnRH, y angiotensina II. Los canales de calcio (cuya compuerta en este caso sería abierta por ligandos intracelulares) se abren indirectamente por acción de los GCPR, a través de la formación de AMPc, GMPc, ATP o el mismo calcio.
Unos más (como el de la somatostatina, receptores alfa 2 adrenérgicos y nor-adrenérgicos) se acoplan con Gi para inhibir adenilciclasa y en algunos tejidos regular los canales de potasio y calcio. Las Go regulan los canales iónicos del sistema nervioso y las Gt (por transducina, que actúa sobre los bastoncitos de la retina), que hidrolizan el GMPc.
Otra forma importante de receptores es la de los tirosina cinasa, que tienen un dominio extracelular para unir los ligandos, un dominio transmembrana y otro citoplasmático, con actividad catalítica. Cuando se fijan hormonas la insulina, el IGF-1, y los factores de crecimiento epidérmico, el derivado de plaquetas y el fibroblástico que utilizan este mecanismo, el receptor se autofosforila al producirse un cambio estructural. La cascada posterior produce la fosforilación de residuos tirosina en otras proteínas.
Como hemos mencionado anteriormente, hay hormonas que utilizan el ion calcio como segundo mensajero, mecanismo en el cual generalmente se encuentran involucrados otros segundos mensajeros como el DAG o el IP3 (fosfoinosítidos), o bien en el aumento en el calcio citosòlico puede intervenir también la apertura de los canales de calcio en la membrana.