I Parte
FELIPE GARCÍA VALLEJO*, MARTHA CECILIA DOMÍNGUEZ**, ADALBERTO SÁNCHEZ***
Introducción
La estructura que conocemos hoy como DNA (ácido desoxirribonucleico) fue descrita por Watson y Crick en el año 1953. A partir de esta propuesta, confluyeron ideas y nuevos enfoques experimentales que condujeron a lo que ha sido denominado como «dogma central de la biología molecular», en el que la información genética fluye del DNA al ARN (ácido ribonucleico) y de este a la proteína; por tanto, propone que es el DNA el material que almacena la información genética en unidades de información hereditaria denominadas genes. Aún cuando este enunciado es cierto para la mayoría de los seres vivientes; se ha comprobado que en determinados virus la información genética se almacena en forma de RNA, esta puede ser transferida del RNA al DNA a través de un proceso de transcripción inversa.
Desde la propuesta de la doble hélice para la molécula del DNA, el conocimiento producido durante más de cuarenta años de investigación en la biología molecular, ha hecho posible el avance de las metodologías que permiten la manipulación del material genético de los organismos, a la vez que ha suscitado una de las mayores revoluciones en el pensamiento biológico como es la de poder entender, de manera más integral, la vida en nuestro planeta. En este contexto, tal vez uno de los mayores logros de la biología molecular en el siglo XX, fue el poder, de manera dirigida, manipular la información genética de los organismos incluyendo el hombre. Este enfoque tecnológico denominado tecnología del DNA recombinante, ha permitido un avance fundamental en la manera como se entiende actualmente el funcionamiento de los genes, en la posibilidad de corregir defectos genéticos y en la producción de nuevas alternativas terapéuticas, todo ello con el fin de mejorar la salud mundial. Sin embargo y a pesar de los grandes avances en estas áreas, existen riesgos potenciales que nos llevan a cuestionarnos cómo estos avances deben utilizarse y regularse para tener una bioseguridad en los protocolos, además de poder darles a todos los habitantes del planeta iguales oportunidades de utilización.
Estructura química del ácido desoxirribonucleico (DNA)
El DNA es un largo polímero (polinucleótido), formado por una serie de subunidades monoméricas denominadas los nucleótidos (figura 1). De acuerdo con el modelo actual más aceptado, el DNA es una doble cadena de polinucleótidos que se enrollan entre si para darle a la molécula de DNA una estructura helicoidal. Una doble cadena de DNA mide de 22 a 26 Å (2.2 a 2.6 nanómetros) de ancho, mientras que (un nucleótido) mide 3.3 Å (0.33 nm) de largo. Aunque cada unidad individual que se repite es muy pequeña, los polímeros de DNA pueden ser moléculas enormes que contienen millones de nucleótidos. Por ejemplo, el cromosoma humano más largo, el cromosoma número 1, tiene aproximadamente 220 millones de pares de bases.
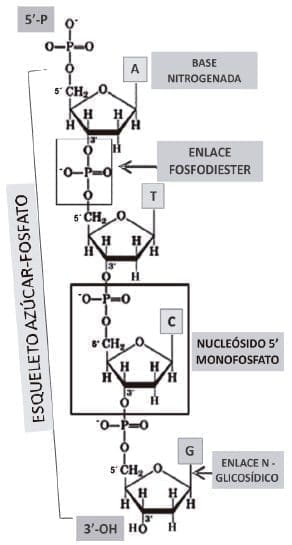 Figura 1. Representación de una cadena polinucleotídica sobre la cual se construyen las moléculas de DNA de doble cadena. Cada uno de los nucleósidos 5´-monofosfato está unido mediante un enlace covalente denominado enlace fosfodiester. Este arreglo hace que la orientación de las desoxirribosas y los fosfatos formen un esqueleto azúcar-fosfato. Este contorno molecular del polinucleótido está cargado negativamente a pHfisiológico y tiene la capacidad de formar enlaces de hidrógeno con le agua. En la otra superficie la interna se dispone las bases nitrogenadas. Así pues el polinucleótido es capaz de tener dos superficies de contacto. El esqueleto azúcar-fosfato es hidrofílico, mientras que la disposición de las distintas bases nitrogenadas forma una superficie relativamente hidrofóbica.
Figura 1. Representación de una cadena polinucleotídica sobre la cual se construyen las moléculas de DNA de doble cadena. Cada uno de los nucleósidos 5´-monofosfato está unido mediante un enlace covalente denominado enlace fosfodiester. Este arreglo hace que la orientación de las desoxirribosas y los fosfatos formen un esqueleto azúcar-fosfato. Este contorno molecular del polinucleótido está cargado negativamente a pHfisiológico y tiene la capacidad de formar enlaces de hidrógeno con le agua. En la otra superficie la interna se dispone las bases nitrogenadas. Así pues el polinucleótido es capaz de tener dos superficies de contacto. El esqueleto azúcar-fosfato es hidrofílico, mientras que la disposición de las distintas bases nitrogenadas forma una superficie relativamente hidrofóbica.
Laboratorio de Biología Molecular y Patogénesis. Departamento de Ciencias Fisiológicas. Escuela de Ciencias Básicas, Facultad de Salud, Universidad del Valle. Correo electrónico: labiomol@gmail.com El éxito del modelo de la doble hélice está en su consistencia con las propiedades físicas y químicas del DNA (figura 2).
La estructura monomérica de soporte de una hebra de DNA es el nucleótido, este a su vez está compuesto de una molécula de azúcar que es una pentosa (la Desoxirribosa); una de cuatro bases nitrogenadas (Adenina, Guanina, Citosina, Timina) cuya unión covalente produce uno de cuatro nucleósidos diferentes. La unión de un grupo fosfato al nucleósido genera el nucleótido. El DNA como polímero está compuesto por largas cadenas de nucleótidos unidas covalentemente entre si por enlaces N-glicosídicos. Por esta razón es que el DNA es una doble cadena de polinucleótidos (figura 1).
La doble hélice del DNA se mantiene estable mediante la formación de puentes de hidrógeno entre las bases asociadas a cada una de las dos hebras. Para la formación de un enlace de hidrógeno una de las bases debe presentar un «donador » de hidrógenos con un átomo de hidrógeno con carga parcial positiva (-NH2 o -NH) y la otra base debe presentar un grupo «aceptor» de hidrógenos con un átomo cargado electronegativamente (C=O o N). Los nucleótidos de cada una de las dos cadenas que forman el DNA establecen una asociación específica mediante puentes de hidrógeno con los correspondientes de la otra cadena. Cada tipo de base en una hebra forma un enlace únicamente con un tipo de base en la otra hebra, lo que se denomina «complementariedad de las bases». Según esto, las purinas forman enlaces con las pirimidinas, de forma que A se enlaza solo con T, y C solo con G.
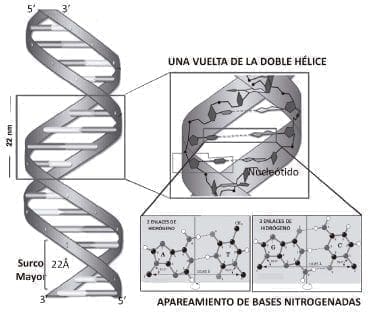 Figura 2. Representación esquemática del modelo de doble hélice para la molécula de DNA. De acuerdo con lo propuesto por Watson y Crick, la doble hélice son dos cadenas polinucleotídicas, cuyas bases nitrogenadas son complementarias, o sea si en una cadena existe A en la complementaria se dispondrá T, estas se parearán mediante dos enlaces de hidrógeno; de otra parte si hay C en la complementaria habrá G y estas dos se unirán mediante tres enlaces de hidrógeno. La doble hélice se favorece termodinámicamente por la sumatoria de esto enlaces de hidrógeno, así la molécula de DNA de doble hélice es una estructura termodinámicamente estable pero flexible. En la doble hélice existirá un surco mayor. Cada vuelta de la doble hélice B incluye 10.5 pares de bases.
Figura 2. Representación esquemática del modelo de doble hélice para la molécula de DNA. De acuerdo con lo propuesto por Watson y Crick, la doble hélice son dos cadenas polinucleotídicas, cuyas bases nitrogenadas son complementarias, o sea si en una cadena existe A en la complementaria se dispondrá T, estas se parearán mediante dos enlaces de hidrógeno; de otra parte si hay C en la complementaria habrá G y estas dos se unirán mediante tres enlaces de hidrógeno. La doble hélice se favorece termodinámicamente por la sumatoria de esto enlaces de hidrógeno, así la molécula de DNA de doble hélice es una estructura termodinámicamente estable pero flexible. En la doble hélice existirá un surco mayor. Cada vuelta de la doble hélice B incluye 10.5 pares de bases.
La organización de dos nucleótidos apareados a lo largo de la doble hélice se denomina apareamiento de bases (figura 2). Este apareamiento corresponde a la observación hecha por Erwin Chargaff quien mostró, que la cantidad molar de adenina era muy similar a la de timina; además que la cantidad molar de citosina era igual a la de guanina en el DNA. Esta observación permitió establecer la hipótesis que una purina siempre mostraba afinidad con una pirimidina.
La doble hélice se estabiliza por el efecto hidrofóbico y el apilamiento de bases nitrogenadas; estos son independientes de la secuencia de bases del DNA. Como los puentes de hidrógeno no son enlaces covalentes, pueden romperse y formarse de nuevo dando una estructura dinámica. Por esta razón las dos hebras de la doble hélice pueden separarse como una cremallera, mediante fuerza mecánica o por alta temperatura. Como resultado de esta complementariedad, toda la información contenida en la secuencia de doble hebra de la hélice de DNA está duplicada en cada hebra, lo cual es fundamental durante el proceso de replicación del DNA. En efecto, esta interacción reversible y específica entre pares de bases complementarias es crítica para todas las funciones del DNA en los organismos vivos.
Existen dos tipos diferentes de apareamiento que ayudan a la estabilización de la doble hélice mediante la sumación de las energías de enlace de los pares de base complementarios. AT forman dos puentes de hidrógeno, y GC tres. El par de bases GC es por tanto, más fuerte que el par de bases AT. Como consecuencia, tanto el porcentaje de pares de bases GC como la longitud total de la doble hélice de ADN determinan la fuerza de la asociación entre las dos hebras de ADN. Así, dobles hélices largas de ADN con alto contenido en GC tienen hebras que interaccionan más fuerte que dobles hélices cortas con alto contenido en AT.
La doble hélice es una espiral dextrógira, esto es, cada una de las cadenas de nucleótidos gira a la derecha. Cuando las dos hebras de ADN se enrollan una sobre la otra, se forman huecos o hendiduras entre ellas, dejando expuestos los laterales de las bases nitrogenadas del interior (figura 2). En la conformación más común que adopta el DNA se generan, como consecuencia de los ángulos formados entre los azúcares de ambas cadenas de cada par de bases nitrogenadas, dos tipos de hendiduras superficiales: El surco mayor, que mide 22 Å (2.2 nm) de ancho, y la otra, la hendidura o surco menor, que mide 12 Å (1.2 nm) de ancho. Cada vuelta de hélice, que es cuando esta ha realizado un giro de 360º mide 34 Å, y en cada una de esas vueltas hay unos 10.5 pares de bases.
Una de las características biológicas de doble hélice es que la secuencia lineal de nucleótidos tiene información biológica. Una proporción de ella está representada en los genes que se codifican por las diferentes proteínas que conforman nuestro cuerpo o hacen parte de los distintos procesos metabólicos. Así pues, la información genética que se transmite a las nuevas generaciones está contenida en la molécula de DNA que en el caso de los humanos, tiene un tamaño de más de 3.000 millones de estos nucleótidos combinados, producto de una evolución biológica.
* Doctor en Ciencias y PDPH. Profesor titular del Departamento de Ciencias Fisiológicas de la Escuela de Ciencias Básicas, Facultad de Salud, Universidad del Valle.
** Magíster en Ciencias con mención en Microbiología. Investigadora en Biología Molecular.
*** PhD y profesor asistente del departamento de Ciencias Fisiológicas de la Escuela de Ciencias Básicas, Facultad de Salud, Universidad del Valle.