Los receptores para las quimioquinas y otros agentes quimioatrayentes pertenecen a la superfamilia de receptores con siete dominios transmembrana tipo serpina o rodopsina, la mayoría de ellos acoplados a proteína G, con tres asas intracelulares y tres extracelulares. Todos presentan un extremo N-terminal extracelular y un C-terminal intracelular. La porción intracelular carboxiterminal contiene residuos serina y treonina, los cuales son fosforilados y participan en la transducción de señales.
La interacción entre la quimioquina y el receptor ocurre en el extremo aminoterminal y una de las asas extracelulares. La proteína G que está acoplada a dicho receptor es heterotrimérica (cadenas a-b-g) y se encuentra unida a la segunda asa intracelular que contiene el motivo DRY (aspartato, arginina, tirosina); dicho motivo ha sido implicado en la transducción de señales.
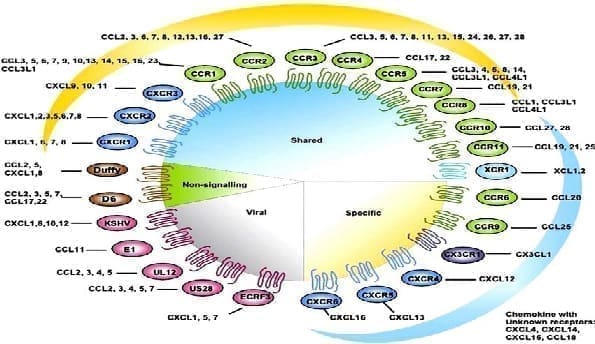
Hasta la fecha, han sido clonados e identificados cinco receptores para la subfamilia CXCL (CXCR1-5) y once para la CCL (CCR1-11). Las quimioquinas muestran gran redundancia en la utilización de sus receptores; de esta manera, varias quimioquinas pueden acoplarse a un mismo receptor y una quimioquina puede ligarse a varios receptores. Por otro lado, los leucocitos, principalmente activados, expresan múltiples receptores debido a la modulación por factores tanto exógenos como endógenos, permitiendo así que en una misma célula la activación de sus receptores por quimioquinas diferentes produzca señales intracelulares distintas. Por lo anterior, se considera que las quimioquinas y otras citoquinas son pleiotrópicas en sus actividades (Tabla 7).
Tabla 7. Receptores para las quimioquinas CXCL: ligandos y expresión celular
Receptor |
Ligandos |
Expresión celular |
| Receptores para CXCL | Neutrófilo: CXCR1, CXCR2 | |
| CXCR1 | CXCL6 (GCP-2), CXCL7 (NAP-2), CXCL8 (IL-8) | Monocito: CXCR4 |
| CXCR2 | CXCL1 (GRO ), CXCL2 (GRO ), CXCL3 (GRO ), CXCL5 (ENA-78), CXCL6 (GCP-2), CXCL7 (NAP-2), CXCL8 (IL-8) |
Linfocito T en reposo: CXCR4Linfocito T activado: CXCR3 |
| CXCR3 | CXCL9 (Mig), CXCL10 (IP-10), CXCL11 (I-TAC) | Linfocito B: CXCR3, CXCR4 |
| CXCR4 | CXCL12 (SDF-1 / ) | C. dendrítica: CXCR4 |
| CXCR5 | CXCL13 (BLC,BCA-1) | Célula NK: CXCR3 |
| Receptores para CCL | ||
| CCR1 | CCL3 (MIP-1 , CCL5 (RANTES), CCL7 (MCP-3), CCL14 (HCC-1), CCL15 (HCC-2), CCL23 (MPIF-1) | Linfocito T activado: CCR1,CCR2, CCR4, CCR5, CCR7 |
| CCR2 | CCL2 (MCP-1), CCL7 (MCP-3), CCL8 (MCP-2), CCL13 (MCP- 4) | Linfocito B: CCR4, CCR5, CCR6, CCR7 |
| CCR3 | CCL5 (RANTES), CCL7 (MCP-3), CCL8 (MCP-2), CCL11 (Eotaxina-1), CCL13 (MCP-4), CCL15 (hcc-1), CCL24 (Eotaxina-2), CCL26 (Eotaxina-3) | Monocito: CCR1, CCR2, CCR5, CCR8 |
| CCR4 | CCL17 (TARC), CCL22 (MDC) | C. dendrítica: CCR1, CCR2, CCR3, CCR4, CCR5, CCR6 |
| CCR5 | CCL3 (MIP-1 , CCL4 (MIP-1 , CCL5 (RANTES) | Célula NK: CCR2, CCR5 Eosinófilo: CCR1, CCR3 Basófilo: CCR2, CCR3 |
| CCR6 | CCL20 (MIP-3 (LARC) | |
| CCR7 | CCL19 (MIP-3 , CCL21 ( 6Ckine, SLC) | |
| CCR8 | CCL1 (I-309), CCL4 (MIP-1 , CCL17 (TARC) | |
| CCR9 | CCL2 (MCP-1), CCL7 (MCP-3), CCL25 (TECK) | |
| CCR10 | CCL27 (CTACK, ILC), CCL28 (MEC) | |
| CCR11 | CCL2 (MCP-1), CCL7 (MCP-3), CCL8 (MCP-2), CCL11(Eotaxina-1), CCL13 (MCP-4) | |
| Receptores para XCLXCR1 | XCL1 (Linfotactina, ATAC) | |
| Receptores para CX3CL | CX3CL1 (Fractalquina) | Linfocito T activado, linfocitos B, monocitos y células NK |
Es importante considerar otro receptor para quimioquinas:
El antígeno Duffy conocido también como DARC; éste tiene la capacidad de unir miembros de las subfamilias CXCL y CCL. La unión ligando-receptor origina la internalización pero no lleva a la transducción de señales; al parecer, el antígeno Duffy tiene un papel regulador similar al que se observa en CCR9, que consiste en impedir el daño inflamatorio mediado por las quimioquinas. Además, se considera útil para el transporte y depuración de éstas en la circulación. Este receptor se expresa en los glóbulos rojos, endotelio de vénulas postcapilares, células de Purkinje (cerebelo) y linfocitos T activados.
Las quimioquinas CXCL ELR+ tienen in vitro acciones redundantes sobre los neutrófilos, incluyendo estímulo de la quimiotaxis, cambios de FORMA, flujos de calcio, degranulación y activación de la explosión respiratoria. Ambos receptores, CXCR1 y CXCR2, son capaces de activar las mismas funciones en los neutrófilos y la vía quimiotáctica independientemente como se ha observado en células Jurkat transfectadas. Sin embargo, estos receptores in vitro parecen ser regulados en FORMA diferente en los neutrófilos humanos, sugiriendo que ellos podrían operar durante diferentes fases de la respuesta inflamatoria. El CXCR2 al parecer tiene un papel clave en el reclutamiento de los neutrófilos, mientras que el CXCR1 es importante en la mediación de señales hacia el sitio de la inflamación, es decir, exocitosis, polimerización de actina y cambios en el pH intracelular.
Funciones proinflamatorias de las quimioquinas
1. Quimiotaxis
Las quimioquinas han sido involucradas en la migración de los leucocitos, tanto en los procesos homeostáticos como inflamatorios.
Las quimioquinas CXCL pueden dividirse de acuerdo a la actividad quimiotáctica en:
1. Proteínas con el motivo ELR (Glu-Leu-Arg) en la región aminoterminal, que son quimiotácticas para neutrófilos y no para macrófagos o monocitos.
2. Sin motivo ELR, son quimiotácticas para células mononucleares y no para neutrófilos.
Las quimioquinas CCL generalmente atraen monocitos y linfocitos, pero no neutrófilos. Son quimiotácticas para los eosinófilos: eotaxina-1, eotaxina-2, MCP-2, MCP-3, MCP-4, RANTES, MDC, MIP-1a (10); algunas de ellas también atraen basófilos. El CCR3 es el receptor que media la mayoría de los efectos quimiotácticos de los eosinófilos y es el único receptor para eotaxina-1 y eotaxina-2.
El rol selectivo de los CCR en la migración de los linfocitos T, monocitos, eosinófilos y basófilos, ha estimulado los ESTUDIOS relacionados con la inflamación alérgica. Muchas quimioquinas tipo CC y CXC regulan la migración de los linfocitos T: IP-10, MIG, RANTES, MIP-1a, MIP-1b, linfotactina. Los linfocitos T vírgenes expresan receptores CXCR4 (para SDF-1) y CCR7 (para ELC), importantes para el tráfico basal de estas células. Las células dendríticas también tienen CCR7; este receptor se utiliza para colocalizar células dendríticas y linfocitos T vírgenes en los nódulos linfoides.
Algunas quimioquinas muestran selectividad por la migración de los linfocitos T activados: MIG, I-TAC, IP-10, 6-C-Kine y STCP-1.
2. Migración transendotelial
ESTUDIOS in vitro han sugerido que la IL-8 es necesaria para la migración transendotelial de los neutrófilos activados; previamente se requiere una señal producida por las citoquinas proinflamatorias IL-1 y TNF-a, que activan las células endoteliales para interactuar con los leucocitos activados por las quimioquinas, en especial la IL-8. El RANTES induce la migración transendotelial de los eosinófilos activados por IL-5.
Las células endoteliales expresan quimioquinas en la superficie luminal, específicamente en los proteoglicanos (glicocálix), estableciendo un gradiente quimiotáctico local; la respuesta migratoria a este estímulo se llama HAPTOTAXIS.
Varias quimioquinas aumentan transitoriamente la adhesividad de las integrinas; por ejemplo, RANTES activa el VLA-4 en los eosinófilos y MCP-3 activa el MAC-1. Otras quimioquinas regulan positivamente la expresión de las integrinas: MIP-1a aumenta la expresión de CD11a y CD11c en monocitos. Algunas quimioquinas pueden funcionar como moléculas de adhesión: la fractalquina y CX3CR1 están en la membrana y regulan el tráfico de monocitos y linfocitos.
3. Activación y degranulación de las células inflamatorias
Referencias bibliográficas
- 1. Springer TA. Traffic signals for lymphocyte recirculation and leukocyte emigration: the multistep paradigm. Cell 1994;76:301-314.
- 2. Zimmerman GA, Prescott SM, McIntyre TM. Endothelial cell interactions with granulocytes: tethering and signaling molecules. Immunol Today 1992;13:93-100.
- 3. Jutila MA. Function and regulation of leukocyte homing receptors. J Leukoc Biol 1994;55:133.
- 4. Tedder TF, Penta AC, Levine HB. Expression of the human leukocyte adhesion molecule, LAM-1: identity with the TQ1 and Leu8 differentiation antigens. J Immunol 1990;144:532-540.
- 5. Jones SL, Lindberg FP, Brown EJ. Phagocytosis. In: Paul WE, ed. Fundamental Immunology. Philadelphia: Lippincott-Raven Publishers, 1999:997-1020.
- 6. Kishimoto TK, Rothlein R. Integrins, ICAMs, and selectins: rolend regulation of adhesion molecules in neutrophil recruitment to inflammatory sites. Adv Pharmacol 1994;25:117.
- 7. Shimizu Y, Newman W, Tanaka Y, Shaw S. Lymphocyte interactions with endothelial cells. Immunol Today 1992;13:106-112.
- 8. Menger MD, Vollmar B. Adhesion melocules as determinants of disease: from molecular biology to surgical research. Br J Surg 1996;83:588-601.
- 9. Carlos TM, Harlan JM. Leukocyte-Endothelial adhesion molecules. Blood 1994;84:2068-2101.
- 10. Chothia C, Jones EY. The molecular structure of cell adhesion molecules. Annu Rev Biochem 1997;66:823-862.
- 11. Horwitz AF. Integrins and Health. Scientifican American 1997;May:46-53.
- 12. Butcher EC, Picker LJ. Lymphocyte homing and homeostasis. Science 1996;272:60-66.
- 13. Moore CM, Ehlayed M, Leiva LE, Sorensen RU. New concepts in the immunolgy of sickle cell disease. Ann Allergy Asth Immunol 1996;76:385-403.
- 14. Passalacqua G, Ciprandi G, Bagnasco M, Battifora M, Canonica CW. Adhesion molecules and allergy – Recent insights. Allergy Clin Immunol International 1998;10:23-29.
Fuentes bibliográficas
- 15. Foster CA. VCAM-1/alpha 4-integrin adhesion patway: Therapeutic target for allergic inflammatory disorders. J Allergy Clin Immunol 1996;98:270-277.
- 16. DeGraba TJ. The role of inflammation after acute stroke: utility of pursuing anti-adhesion molecule therapy. Neurology 1998;51:62-68.
- 17. Nickel R, Beck LA, Stellato C, Schleimer RP. Chemokines and allergic disease. J Allergy Clin Immunol 1999;104:723-742.
18. Barrett JR. Chemokines. Blood 1997;90:909-928. - 19. Ben-Baruch A, Michiel DF, Oppenheim JJ. Signals and receptors involved in recruitment of inflammatory cells. J Biol Chem 1995;270:11703-11706.
- 20. Baggiolini M, Dewald B, Moser B. Human chemokines: an update. Annu Rev Immunol 1997;15:675-705.
- 21. Alam R. Chemokines in allergic inflammation. J Allergy Clin Immunol 1997;99:273-277.
- 22. Luster AD. Chemokines: chemotactic cytokines that mediate inflammation. New England J Med 1998;338:436-445.