La lactancia es fundamental
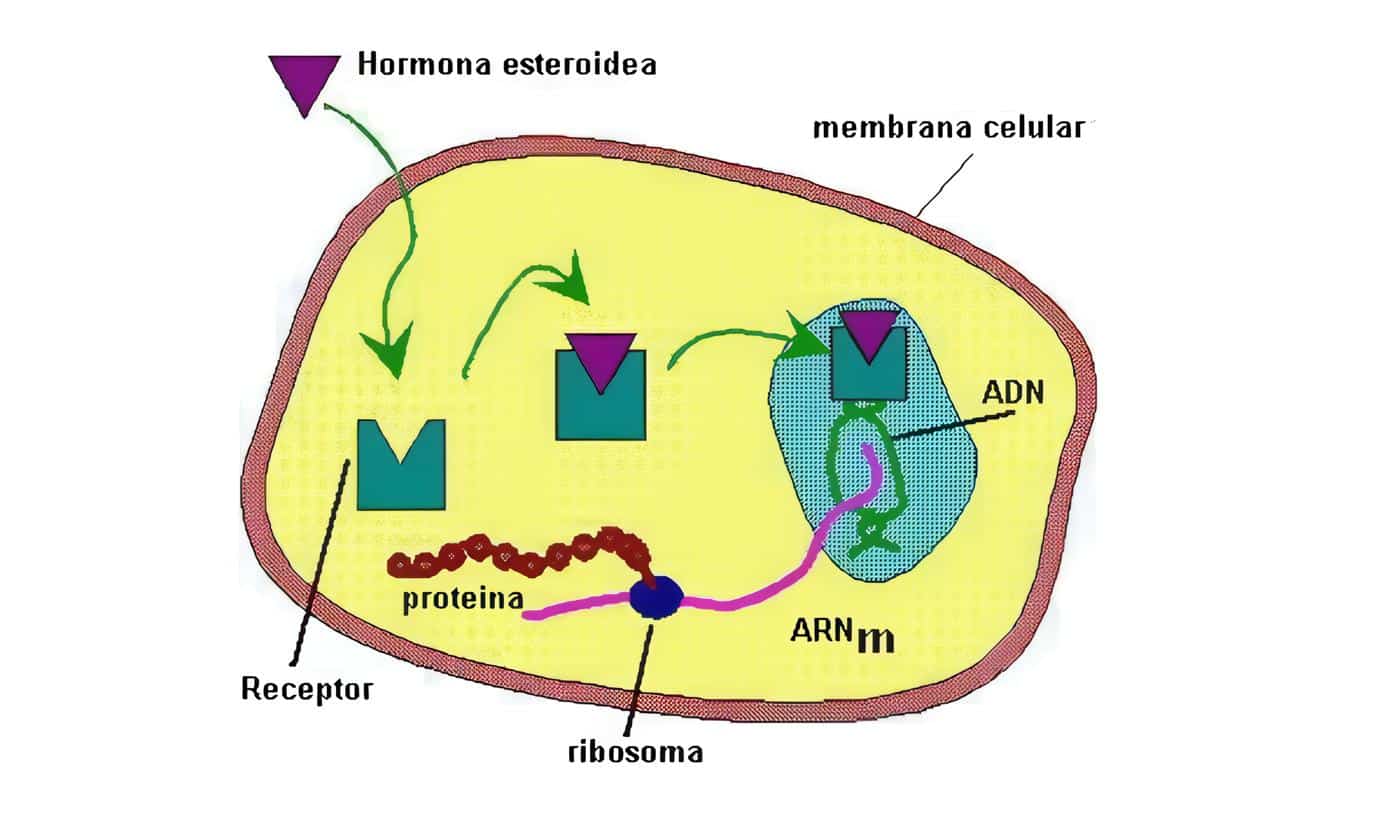
La lactancia es fundamental para la conservación de la especie, ya que de ella depende que el recién nacido reciba la cantidad necesaria de nutrientes para poder continuar con su crecimiento y desarrollo. Parte de los requisitos necesarios para garantizar una lactancia adecuada dependen de la producción hormonal, especialmente en lo que tiene que ver con prolactina.
Para que la lactancia pueda ser iniciada se hace indispensable el crecimiento del tejido mamario, hecho que ha recibido el apelativo de mamogénesis. Durante la gestación las células epiteliales secretoras pasan por un período de diferenciación, conocido como lactogénesis I. El proceso culmina después del parto, cuando aparece la producción abundante de leche, llamada lactogénesis II; este último paso se encuentra directamente ligado a los diferentes procesos que gobiernan el comienzo del trabajo de parto.
Desde las primeras semanas de gestación el seno empieza a crecer y prepararse para la lactancia, gracias a los cambios hormonales observados, especialmente la elevación de la prolactina.
Esta hormona es producida por los lactotropos de la hipófisis anterior.
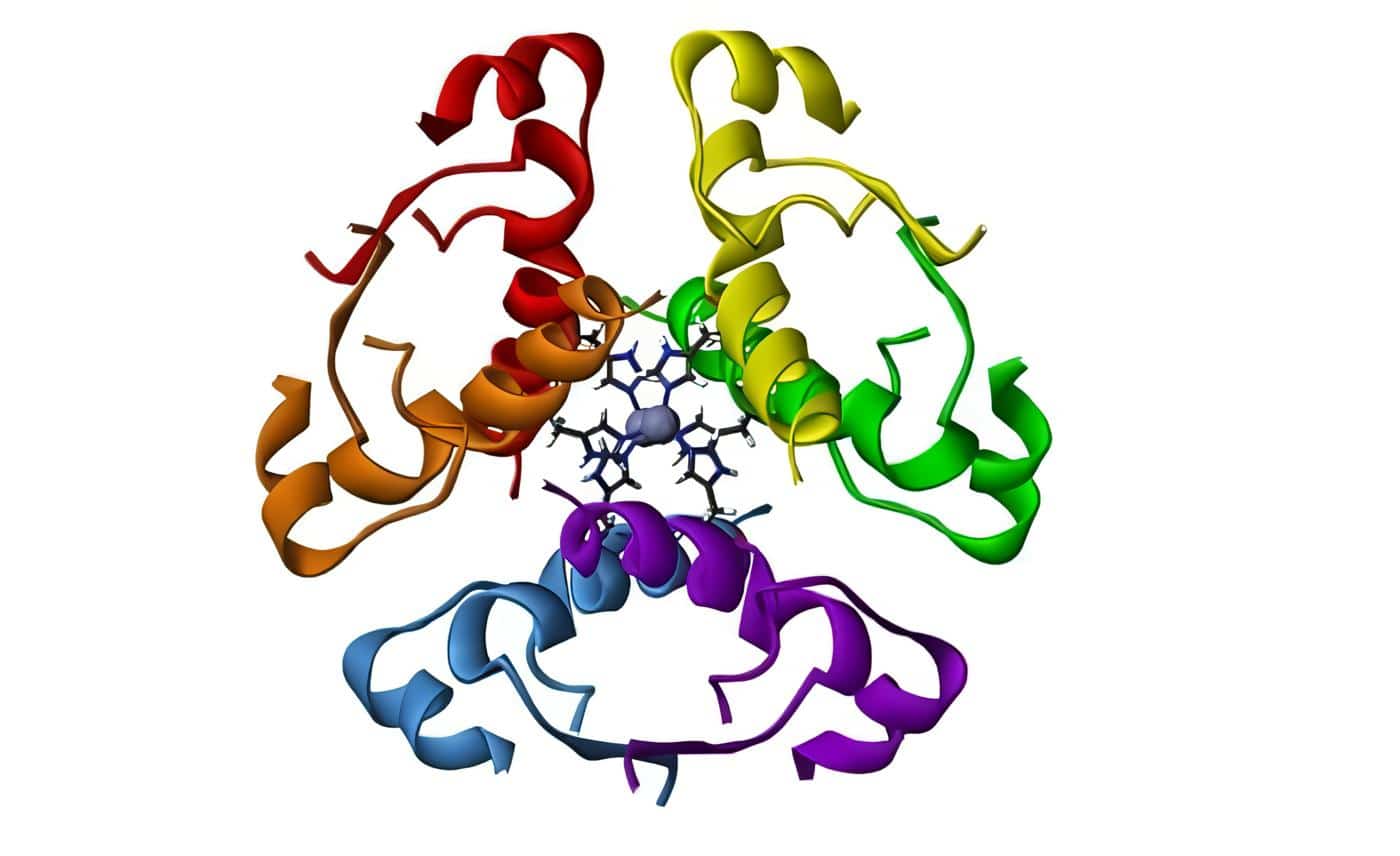
La prolactina es un polipéptido que contiene 198 residuos de aminoácidos con un peso molecular de aproximadamente 22.000. Su estructura está plegada para adoptar una forma globular y los pliegues están conectados por tres uniones disulfuro. Su secuencia es homóloga a la de la hormona de crecimiento y al lactógeno placentario. Durante el embarazo aumenta la prolactina circulante y en forma simultánea el número y el tamaño de las células lactotropas en la hipófisis anterior. Este aumento de la función lactotropa se produce como resultado del incremento en los niveles de estrógenos. En el feto la prolactina plasmática sigue cambios similares paralelos a los niveles de estrógenos. Los estrógenos intervienen en el metabolismo de la dopamina, de modo que el tono dopaminérgico se encuentra disminuido en la eminencia media. Existe un efecto directo de los estrógenos sobre la célula lactotropa incrementando su capacidad de síntesis y su respuesta a los estímulos liberadores. Es por ello que los niveles séricos ascienden progresivamente hasta alcanzar valores que oscilan entre 200 y 400 ng/ml. Gracias a este efecto de la prolactina hay un incremento de los racimos alveolares, los alvéolos diferenciados se vuelven más evidentes y hay hipertrofia celular.
Durante la lactogénesis I se adquiere la posibilidad de empezar a secretar sustancias de la leche como:
Lactosa, caseína y lactalbúmina, las cuales son nuevamente reabsorbidas. Este cambio en la glándula mamaria puede observarse entre las 10 y las 22 semanas de embarazo y es secundario a la presencia de prolactina.
Durante el embarazo los niveles de prolactina aumentan desde su valor normal hasta 200 o 400 ng/ml al término; este aumento empieza hacia las 8 semanas en forma simultánea con el incremento en los estrógenos. Se cree que el aumento en la secreción de prolactina es debido a la supresión que los estrógenos producen sobre la dopamina y por estimulación directa de la transcripción del gen de la prolactina en la hipófisis.
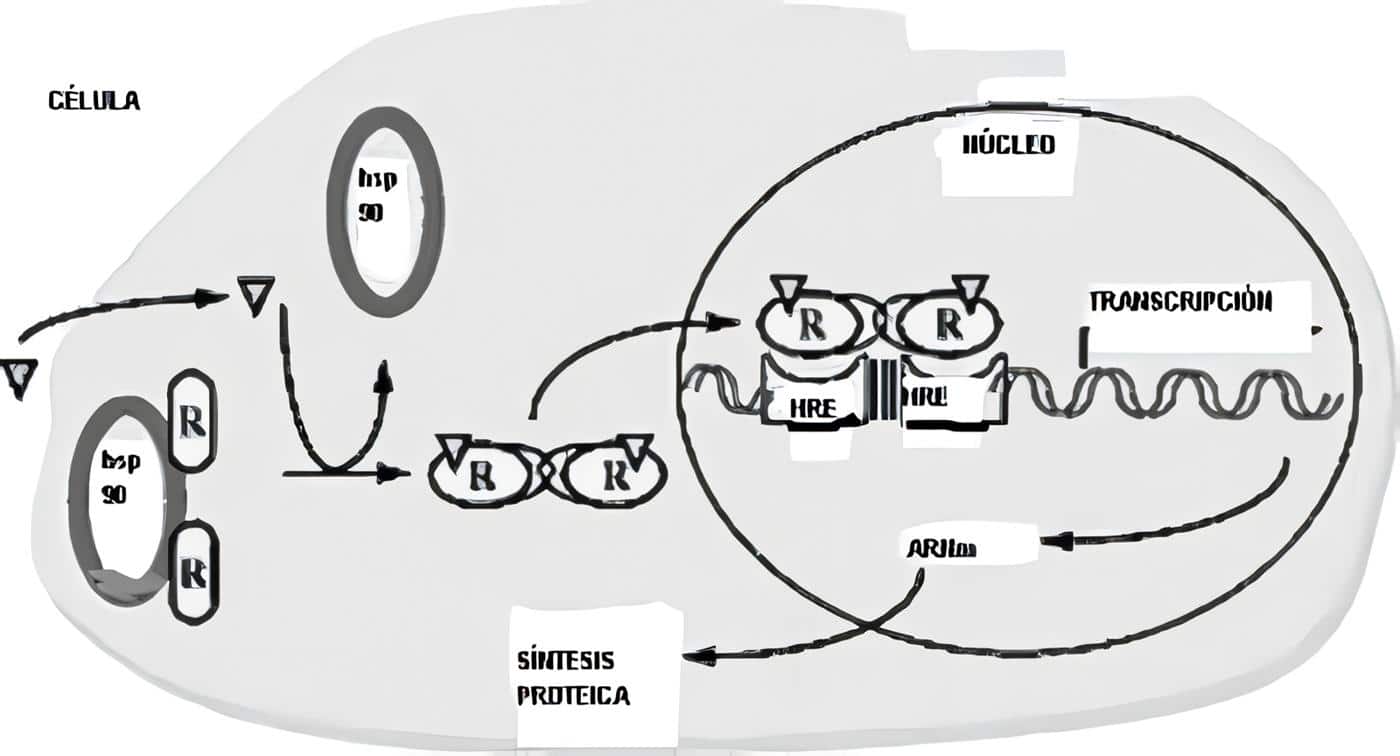
Aunque la prolactina estimula el crecimiento del seno y está disponible para iniciar la lactancia, durante el embarazo solo se produce calostro compuesto por células epiteliales descamadas. La lactancia como tal se encuentra inhibida por la progesterona que interfiere con la acción de la prolactina sobre su receptor. Tanto los estrógenos como la progesterona son necesarios para la expresión del receptor lactogénico. Se cree que los niveles elevados de estrógenos pueden bloquear la acción de la prolactina. Al parecer el IGF-I puede jugar algún papel a través de la inducción en la síntesis de caseína.
En ausencia de la prolactina no hay síntesis de caseína, proteína indispensable en la leche.
El gatillo que dispara la producción de leche dentro de la célula alveolar es la supresión rápida de los estrógenos y progesterona después del parto, llevando a la aparición de la lactogénesis II. La depuración de prolactina es mucho más corta en este momento, necesitando hasta 7 días para llegar a los niveles previos al embarazo cuando no hay lactancia. En los casos en los cuales hay lactancia, en la primera semana postparto los niveles disminuyen más o menos a la mitad. La succión produce un aumento en la concentración de prolactina, estímulo indispensable para iniciar la producción de leche. Dos a tres meses postparto los valores basales pueden estar alrededor de 40 o 50 ng/ml, con aumentos de 10 a 20 veces después de la succión.
El mantenimiento de la producción de leche en niveles elevados es dependiente de la acción conjunta de la hipófisis anterior y la posterior; la succión induce la liberación de prolactina, oxitocina y TSH. La prolactina a su vez sostiene la síntesis de caseína, ácidos grasos y lactosa, además de mantener el volumen de secreción; la oxitocina hace contraer las células mioepiteliales y desocupa la luz alveolar, estimulando una mayor secreción de leche. El aumento de la TSH sugiere que la TRH puede jugar papel en la respuesta de la prolactina a la succión.
Las mujeres que lactan en forma regular, incluyendo la noche, tienen una protección anticonceptiva por tres a seis meses.
Cuando están aumentados los niveles de prolactina por el efecto de la succión, se encuentran niveles de FSH en el rango normal pero abolición de los picos de LH. La secreción de estrógenos en el aparato folicular es muy baja. El efecto puede no solo ser central sino ovárico, afectando la función de las células de la granulosa disminuyendo la síntesis de progesterona. También puede alterar la relación testosterona/dihidrotestosterona, disminuyendo el sustrato androgénico para la aromatización. En todo caso, parece que la acción central es la más importante; los niveles elevados de prolactina inhiben la secreción pulsátil de GnRH, por el asa corta de retroalimentación sobre la dopamina.
Las concentraciones de prolactina en el líquido amniótico aumentan en forma paralela a los niveles sanguíneos maternos hasta la décima semana de embarazo; se incrementan en forma marcada hasta la semana 20 y luego disminuyen. La prolactina materna no pasa al feto en cantidad significativa.
Esta prolactina del líquido amniótico es un producto decidual, transferido por receptores en el amnios hacia el líquido amniótico, para lo cual se requiere la integridad del amnios, el corion y la decidua. La síntesis decidual de prolactina es iniciada por la progesterona, pero una vez que se ha establecido la decidualización, su síntesis continúa incluso en ausencia de los esteroides sexuales. Hay varios factores deciduales comprometidos en la regulación de la prolactina, incluyendo la relaxina, la insulina y el IGF-I. Se postula que el papel que juega la prolactina en el líquido amniótico es regular el transporte de sodio y agua evitando la deshidratación del feto.
La prolactina interviene en muchos eventos bioquímicos durante el embarazo.
En el feto influye sobre la síntesis del surfactante pulmonar y la prolactina decidual modula la contractilidad del músculo liso uterino mediada por prostaglandinas. También contribuye a evitar el rechazo inmunológico del concepto, suprimiendo la respuesta inmune materna.
Resumen
Este capitulo se refiere a los cambios hormonales que sufre la mujer durante el embarazo, donde hay un medio interno con niveles hormonales muy altos, inicialmente dirgidos desde el cuerpo lúteo por acción de la progesterona, y luego por la gonadotropina coriónica. Se discuten también los mecanismos y cambios hormonales durante el trabajo de parto y posparto, además de los efectos hormonales sobre la glándula mamaria y la lactancia. En cuanto a esto último, los estrógenos aumentan el tejido adiposo, la progesterona los alvéolos secretores, la prolactina la producción de leche materna y la ocitocina es la hormona eyecto-láctea.
Referencias
- Barón G. Fisiología del ciclo menstrual. Endocrinología del embarazo. 1era. Edición. Colombia: Editorial Contacto Gráfico: 1998.
- Barón G. Fisiología del ciclo menstrual. Hiperprolactinemia. 1era. Edición. Colombia: Editorial Contacto Gráfico: 1998.
- Abe Y, Hasegawa Y, Miyamoto K, et al. High concentrations of plasma immunoreactive inhibin during normal pregnancy in women. J Clin Endocrinol Metab 1990;71:133.
- Albrecht ED, Pepe GJ. Placental steroid hormone byosynthesis in primate pregnancy. Endocrin Rev 1990;11:124
- . Begum-Hasan J, Murphy BEP. In vitro stimulation of placental progesterone production by 19-nortestosterone and C-19 steroids in early human pregnancy. J Clin Endocrinol Metab 1992;75:838.
- Emmi AM, Skurnick J, Goldsmith LT, et al. Ovarian control of pituitary hormone secretion in early human pregnancy. J Clin Endocrinol Metab 1991;72:1359.
- Handwerger S. Clinical counterpoint: the physiology of placental lactogen in human pregnancy. Endocr Rev 1991;12:329.
- Handwerger S, Brar A. Placental lactogen, placental growth hormone, and decidual prolactin. Seminars Reprod Endocrinol 1992;10:106.
- Jameson JL, Hollenberg AN. Regulation of chorionic gonadotropin gene expression. Endocr Rev 1993;14:203.
- Kliman HJ. Placental hormones. Infertil Reprod Med Clin of North Am 1994;5:591.
- Petraglia F, Woodruff TK, Botticelli G, et al. Gonadotropin-releasing hormone, inhibin, and activin in human placenta: evidence for a common cellular localization. J Clin Endocrinol Metab 1992;74:1184.
- Reis FM, Florio P, Cobellis L, et al. Human placenta as a source of neuroendocrine factors. Biol Neonate 2001;79:150-6.
- Malassiné A, Frendo JL, Evain-Brion D. A comparison of placental development and endocrine functiones between the human and mouse model. Hum Reprod Update 2003;9:531-9.
Bibliografías
- Forsyth IA, Wallis M. Growth hormone and prolactin – molecular and functional evolution. J Mammary Gland Biology and Neoplasia 2002;7:291-312.
- Kleine B, Wolfahrt S, Lotsch M, et al. Expression of galanin in human placenta. Mol Hum Reprod 2001;7:379-85.
- Carbillon L, Uzan M, Challier JC, et al. Fetal-placental and decidual placentalunits: role of endocrine and paracrine regulations in parturition. Fetal Diagn Ther2000;15:308-18.
- Battin DA, Marrs RP, Fleiss PM, et al. Effect of sucking on serum prolactin, luteinizing hormone, follicle-stimulating hormone, and estradiol during prolonged lactation. Obstet Gynecol 1985;65:785.
- De Greef WJ, Voogt JL, Visser TJ, et al. Control of prolactin release induced by suckling. Endocrinology 1987;121:316.
- Liu JH, Lee DW, Markoff E. Differential release of prolactin variants in postpartun and early follicular phase women. J Clin Endocrinol Metab 1990;71:605.
- Hartmann PE, Cregan MD, Ramsay DT, et al. Physiology of lactation in preterm mothers: initiation and maintenance. Pediatric Ann 2003;32:351.
* El profesor Germán Barón Castañeda es Médico Ginecólogo Endocrinólogo de la Universidad del Rosario, Profesor Distinguido de la Universidad del Rosario y Profesor de la Universidad del Bosque. Jefe de Educación Médica del Hospital Universitario de la Samaritana. Es autor del libro FUNDAMENTOS DE ENDOCRINOLOGÍA GINECOLÓGICA.
Dirigió por diez años la sección de endocrinología ginecológica del Hospital Infantil Lorencita Villegas de Santos en Bogotá, Colombia. Estos dos artículos hacen parte del libro Fisiología Endocrina, Tercera Edición, cuyo autor principal es Alfredo Jácome Roca, y que se publicará próximamente.